

par Pierluigi Angeli et Maria Tullii

Traduction en français de Bruno Scaramella
QUELQUES ÉTAPES IMPORTANTES DE L’HISTOIRE DE LA MYCOLOGIE

Agaricus praeclaresquamosus © Giuseppe Mazza
La mycologie est la science qui étudie la branche de la botanique qui s’intéresse aux champignons. Le terme MYCOLOGIE tient ses origines du mot grec MÝCHÊS = champignon et de LÒGOS = discours, traité. Cela implique que mycologie veut dire, littéralement, avoir affaire aux champignons. Un mycologue est une personne qui étudie les champignons.
Pour pouvoir débattre de champignons ainsi que pour comprendre certaines coutumes et attitudes, il est parfois utile de remonter le temps très loin et même jusqu’à la préhistoire.
Sur la base des témoignages recueillis (graffitis préhistoriques), on peut supposer que l’homme a commencé à observer les champignons à la nuit des temps. Il a d’abord appris à s’en servir come nourriture, tout en découvrant leur dangerosité à ses risques et périls puis, après avoir décelé les propriétés particulières de certaines variétés de champignons, hallucinogènes pour certains, dont se servaient notamment les sorciers et les chamans, l’homme a également pu bénéficier des qualités curatives de certaines espèces, notamment de leurs propriétés anti-inflammatoires et de renfort de nos défenses immunitaires. Aussi, à des époques relativement plus récentes, les champignons ont été employés pour éliminer les ennemis et les personnes dites “gênantes”.
Il est facile d’imaginer comment, au début, les empoisonnements causés par la consommation de champignons pouvaient être fréquents. Les difficultés éprouvées pour comprendre les causes d’intoxication firent naître des superstitions et des fausses croyances qui subsistent à bien d’endroits encore aujourd’hui.

Amanita pantherina © Giuseppe Mazza
On pensait que les champignons, inoffensifs à la base, pouvaient devenir toxiques par l’intervention de facteurs extérieurs ou alors, qu’ils étaient une création du démon pour attirer l’homme et lui faire du mal !
Cette manière de concevoir les champignons est probablement due à son mode de naissance et de croissance : il émerge de la terre apparemment de rien (cette “plante-champignon” se développe sous terre, ne produit pas de “semences” dans le vrai sens du terme et la croissance de ses sporophores n’est pas liée à un cycle saisonnier particulier mais plutôt aux conditions climatiques qui varient d’une année sur l’autre) ; de plus, la croissance des sporophores, prenant souvent la forme d’un cercle ou de lignes zigzagantes, était interprétée comme une manifestation du mal.
Les preuves qu’on se servait de certaines espèces pour provoquer des hallucinations et des états de transes lors de certains rites chamanistes ou divinatoires, sont contenues dans les nombreuses peintures rupestres et les dessins retrouvés à plusieurs endroits du monde : le désert du Sahara, Alger, la Vallée des Merveilles en France, l’Inde, le nord de l’Europe ; partout, ces représentations suggèrent la consommation du genre Psilocybe ou alors de champignons qui décèlent des propriétés hallucinogènes au moins.
L’utilisation divinatoire n’était toutefois pas le seul emploi réservé aux champignons et il en existe un témoignage récent. Le corps naturellement momifié d’un homme, qui a vécu il y a 5 000 ans, a été retrouvé au niveau du glacier du Similaun, qui sépare l’Italie de l’Autriche, en 1991.

Le Fomes fomentarius était utilisé dans l’antiquité pour faire du feu © Pierluigi Angeli
Le Fomes fomentarius et le Lenzites betulinus faisaient partie des objets qui constituaient son équipement ; ce premier servait à allumer un feu alors que le deuxième était utilisé pour accroître la résistance physique à la fatigue et à la douleur de l’homme lorsqu’il était absorbé sous forme d’infusion, ou bien comme antibiotique et hémostatique pour traiter les blessures sous forme de poudre et probablement aussi comme brosse pour déterger la peau. Encore aujourd’hui, le Lenzites betulinus, qui est considéré un tonique naturel dans les pays scandinaves, est pris sous sa forme infuse. Même s’ils ne sont guère nombreux, des témoignages écrits d’époques plus récentes sur les champignons, leur nature, leur utilisation, ainsi que les croyances auxquelles ils sont liés, existent aussi.
Un des textes documentés sur les champignons les plus anciens fait partie d’une œuvre de la Grèce Antique et traite de la manière dont ils sont cultivés : “ …enterrer du fumier de cheval sous un figuier et l’arroser abondamment… ”
Le grec THÉOPHRASTE (370-287 av. J.-C) natif de l’île de Lesbos et disciple d’ARISTOTE, qui est considéré comme le père de la botanique s’exprime ainsi au sujet des champignons : “…ce sont des plantes imparfaites, dépourvues de racines, de feuilles, de fleurs et de fruits”.
Après THÉOPHRASTE, le domaine des champignons devint l’apanage des médecins et il faudra attendre l’année 50 de notre ère pour lire d’autres textes sur ceux-ci. PEDANIUS DIOSCORIDE, médecin militaire et civil, né en Cilicie (Asie Mineure), au service de Rome sous les empereurs Claudius et Néron, nous a transmis par son ouvrage “DE MATERIA MEDICA” (Sur les Plantes Médicinales) des notions sur la toxicité des champignons et sur les thérapies qu’il faut suivre en cas d’empoisonnement : “des infusions d’herbes aromatiques, des potions de vinaigre et de sel, l’administration de déjections de poulet empâtées avec du miel et du vinaigre…”, des remèdes qui ont probablement été utilisés pendant des siècles.

Boletus satanas © Giuseppe Mazza
C’est à lui qu’on doit aussi la première description de l’AGARICUM (Laricifomes officinalis) qui était utilisé comme médicament.
Un autre docteur grec de Pergame, CLAUDE GALIEN (129-200 ap. J.-C), qui vécut à Rome aux temps des empereurs Marc Aurèle, Lucius Verus et Commode, s’est intéressé aux champignons également. Galien a subdivisé les champignons en trois groupes principaux : “Bolites”, “Amanitai” et “Mikés”.
Le premier groupe regroupait les champignons à lames (dont fait partie l’Amanite actuelle qui est un genre auquel appartient l’ Amanita caesarea qu’on appelle couramment “ bolet ” dans certaine régions italiennes) ; le deuxième groupe englobait tous les champignons de forme tubulaire ou poreuse (tel le genre Boletus selon l’appellation qui lui a été attribuée par Linnæus en 1735) ; le troisième groupe incluait enfin tous les autres champignons à chapeau et tige.
Les champignons appartenant aux deux premiers groupes, devaient tous être considérés comme comestibles, selon Galien, alors que ceux qui faisaient partie du troisième groupe devaient être jugés toxiques et non adaptés à la consommation car : “…non nutritifs, à la digestion difficile et dangereux de par leurs propriétés toxiques…”. Galien confirmât aussi la validité de la thérapie à base d’excréments de poulet comme remède contre les empoisonnements.
Gaius Plinius Secundus, connu sous le nom de PLINE L’ANCIEN (23-79 ap. J.-C.), est le premier auteur à avoir traité les champignons d’une manière plus approfondie. Il nous a légué une œuvre de 37 livres : “HISTORIA NATURALIS” où, parmi les autres sujets naturalistes, il décrit aussi les champignons. Citons-en quelques passages : “…nous estimons devoir inclure les bolets parmi les aliments les moins conseillés…” (ndlr les Amanites actuelles) et il justifie cette affirmation par la facilité de confusion avec les espèces toxiques.

Amanita muscaria © Giuseppe Mazza
En ce qui concerne l’ Amanita muscaria, il écrit : “…certains se distinguent par de sortes de verrues blanches et sèches, semblables à une efflorescence de salpêtre, qui ne sont autre que des résidus du voile, présents sur leur chapeau… ”.
Ensuite, Pline donne des exemples de circonstances dans lesquelles des champignons parfaitement mangeables peuvent devenir toxiques en disant : “…s’ils naissent près de boutons métalliques, de clous de chaussure, de morceaux de fer rouillés, de chiffons détrempés, ils deviennent empoisonnés en absorbant les sucs imprégnés de ces substances”.
Et encore : “…s’il y a un nid de serpents dans les parages, ou bien si un serpent souffle sur le champignon lors de son passage, il devient alors empoisonné, car sa nature est celle d’absorber toute sorte de substance vénéneuse”.
De telles croyances sont encore omniprésentes et jugées comme véridiques sur la quasi-totalité du territoire italien.
On pensait aussi pouvoir déterminer si un champignon était ou non comestible en observant les plantes qui poussaient près de lui : Pline jugeait que les champignons qui naissent sous les conifères, les figuiers et les férules sont inoffensifs alors que ceux qui apparaissent sous les chênes, les hêtres et les cyprès sont vénéneux. Ceci est probablement dû au fait que les Amanita mortelles et la phalloides privilégient particulièrement les arbres feuillus, alors que nous savons que le Pleurotus eryngii, qui est un champignon délicieux connu depuis l’antiquité, est issu des racines de la férule.
Pline décrit également le développement des bolets : “…ils naissent de la terre, d’abord enveloppés de leur volve qui leur donne l’apparence d’un œuf, ce qui fait qu’ils soient particulièrement appréciés à ce stade de la croissance. Ensuite, la volve se brise en restant accrochée à la base de la tige… ”

Amanita phalloides © Giuseppe Mazza
C’est la première fois que le terme volve est employé dans son vrai sens mycologique. Il décrit ensuite la durée de vie d’un bolet qui est de sept jours à peine ainsi que sa croissance cespiteuse avec ses sporophores non couplés mais individuels.
Afin d’expliquer la nature des champignons (qui apparemment naissent du néant) Pline dit : “…il faut rechercher leur origine dans le limon de la terre humide et dans son humus en phase de fermentation ou bien dans les racines des plantes cupulaires…”
Cette affirmation demeurera enracinée dans les opinions des naturalistes pendant des siècles jusqu’à ce qu’on puisse donner une connotation à l’étude de la matière par le biais d’instruments modernes.
Pline offre une place de choix au bolet en le décrivant à part alors qu’il groupe ensemble tous les autres champignons en affirmant : “…les autres champignons ont une consistance plus ferme. Bien qu’il existe une infinité de types, ils possèdent tous une origine commune : la sève des arbres. Ceux qui ont une couleur rouge plus foncée que les bolets sont certainement comestibles… ”.
C’est sans doute aux champignons lignatiles et certainement à la Fistulina epatica, qui était considéré comme très appétissant dès l’antiquité, que faisait allusion Pline.
Il poursuit son discours : “…il y a ensuite des espèces de couleur blanche à la tige très développée et au chapeau qui ressemble un couvre-chef d’un Flamine… ” Le Flamine était un prêtre d’une religion romaine ancienne qui portait un chapeau dont l’aspect suggérait un Macrolepiota procera au chapeau seulement partiellement ouvert.
Et : “…un troisième type de champignon, le Suilli a un gros penchant pour le poison…”

Fistulina hepatica © Giuseppe Mazza
Les Suilli étaient ce qu’on appelle à l’heure actuelle les Boletaceae. On les considérait vénéneux à cause de leur couleur livide, c’est-à-dire bleuâtre.
Par contre les “Suilli” étaient un produit très commercialisé au temps de Pline : ils étaient importés d’Asie Mineure et on les préparait en les embrochant à des joncs très fins pour qu’ils sèchent près du feu. Ce système est utilisé encore de nos jours dans certaines régions italiennes.
Certains Suilli étaient également utilisés par les médecins pour soigner les hémorroïdes, les boutons, les morsures et même dans la préparation des collyres. Toutefois, Pline estimait que le danger majeur venait des porcins toxiques qui sont responsables d’empoisonnements de masse au sein de moult familles.
Dans son traité il conseille : “…si quelqu’un veut vraiment les consommer, qu’il fasse très attention au moment de leur achat et qu’il soit très attentif à leur cuisson…”
Et de conclure : “…qu’il se méfie des champignons qui durcissent à la cuisson ; qu’il les fasse cuire longuement” en ajoutant du salpêtre à la poêle. Qu’on les cuise de préférence avec de la viande (pour les diluer) avec des pétioles de poires sauvages (autrement ces derniers pourront être servis à la fin du repas en tant qu’antidote). Qu’on se souvienne également que le vinaigre est un antidote naturel et qu’il fasse donc l’objet d’une utilisation abondante lorsqu’on mange des champignons”
Il faut remarquer que si on suit ce dernier conseil lors de la préparation du Coprinus atramentarius on risque une intoxication très grave comme un syndrome coprinien.

Macrolepiota procera © Giuseppe Mazza
DU MOYEN-AGE AU XVIII SIECLE
Avec la chute de l’empire romain d’occident et la barbarisation des peuples à cause des invasions des peuples nordiques, on entre dans ce qui est connu sous le nom de “moyen-âge”. La culture classique et scientifique passe dans les mains du clergé et surtout dans celle des moines Bénédictins qui s’occupent alors de la conservation des œuvres, de leur transcription et de leur traduction.
Nous sommes assez pauvres en informations concernant le moyen-âge et rares sont les érudits en matière de champignons qui nous ont transmis des œuvres. La Renaissance, par contre, voit le commencement d’une nouvelle ère pour la botanique, qu’on reconnaît désormais comme une vraie science même si elle est complémentaire de la médecine. Cette période connaît aussi la révision des œuvres de Dioscoride et de Pline avec des traductions, des commentaires et des rééditions.
HERMOLAO BARBARO (1454-1492) amateur de la culture d’Aristote, de Pline et de Dioscoride, a traduit et commenté leurs œuvres.
PIETRO ANDREA MATTHIOLI (1500-1577) né à Sienne mais originaire de Trente, était un médecin notoire. Commentari in Pedacii Dioscoridis Anarzabei de Materia medica, son œuvre première, contient la traduction de l’œuvre originale de Dioscoride du grec au Latin. A chaque chapitre correspond une annexe avec un commentaire qui est encore plus long que le texte du chapitre lui-même. L’œuvre de Matthioli était très prestigieuse au point de connaître 60 éditions sur deux siècles dans sa langue d’origine, en français, en allemand et en tchèque. Bien que la communauté scientifique eût déjà considéré la botanique comme une vraie discipline, toujours en tant que branche de la médecine, on ne s’occupait que marginalement des champignons. Le seizième siècle vit la naissance des premiers traités monographiques. Le premier d’entre eux fut publié par un médecin hollandais JONGHE HADRIAN (1512-1575) lequel composa une dissertation au sujet d’une nouvelle espèce qu’il nomma Phallus impudicus, sous le titre “Phalli ex fungorum genere in Hollandiae sabuletis passim crescentis, descripto et ad vivum expressa pictura”, où un champignon était décrit et illustré à l’aide d’une table xylographique, dont le nom sera changé plus tard en Phallus hadriani en son honneur.

Rhizomorphes au niveau du sous-sol © Giuseppe Mazza
Ce champignon poussait sur les terres sableuses de Hollande et le fascicule qui lui était dédié rapportait également ses qualités médicamenteuses qui le faisaient considérer comme thérapeutique pour le traitement de la goutte et les douleurs arthritiques.
ANDRE CÉSALPIN (1525-1603) né à Arezzo en 1525 et presque contemporain de Matthioli, un médecin tout aussi célèbre, a laissé une empreinte indélébile dans le milieu de la botanique. Dans le domaine médical, il fut le premier à découvrir et à décrire en des termes modernes le système de circulation du sang et les capillaires même si le mérite revint injustement, plus tard, à l’anglais Harvey. Césalpin fut le médecin particulier du Pape Clément VIII, archiatre pontifical et professeur de médecine à l’université “la Sapienza” de Rome. Il fut aussi le médecin de Saint Philippe Néri et du Tasse. Son œuvre en 16 volumes De Plantis, est considérée sa contribution principale au domaine de la botanique.
C’est la toute première tentative de cataloguer les plantes de manière systématique et selon des critères précis. On le considère pour cette raison le père de la botanique italienne. Césalpin, en tant que botaniste, traite aussi les champignons bien sûr ; il est le premier à s’essayer à une étude approfondie de ceux-ci et il est également le premier à fournir, en plus de la description des différentes espèces, des conseils sur la cuisson des champignons comestibles. On ne peut pas dire toutefois que la connaissance des champignons avait fait des pas de géant jusque là.
L’influence des croyances anciennes pesait toujours, l’absence d’instrumentation scientifique adaptée laissait des vides surtout en ce qui concernait leur manière de naître, on pensait que les champignons étaient des manifestations pathologiques des racines ou des arbres, quand on ne leur attribuait pas des origines magiques. Alors que le progrès de la botanique a été remarquable, on ne peut en dire autant de la connaissance des champignons.
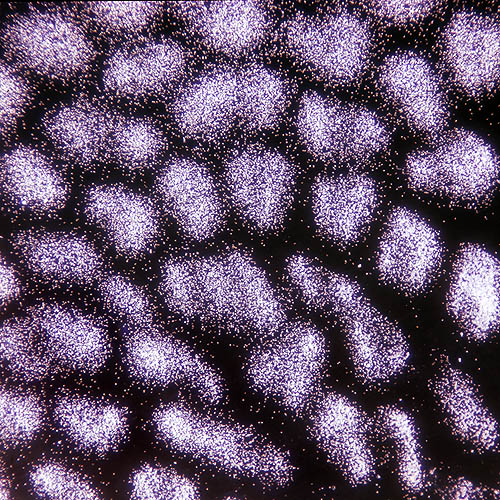
Spores de Boletus aestivalis vues au microscope à faible agrandissement © G. Mazza
DU XVIII SIECLE A NOS JOURS
Il y a eu plusieurs savants qui se sont dédiés aux champignons et qui ont essayé de les classer dans le domaine de la botanique. Toutefois, ce n’est qu’avec l’arrivée de PIETRO ANTONIO MICHELI (Florence 1679-1737) que la mycologie moderne vient à naître ainsi que nous la connaissons encore aujourd’hui.
Micheli développe sa passion pour la botanique en lisant les œuvres de Matthioli, de Bocconi et d’autres savants alors qu’il travaille en tant que garçon de boutique chez un relieur de livres. Il devient un botaniste de renommée internationale, poussé par sa grande passion, même s’il n’a obtenu aucun diplôme, aidé et éduqué par des moines Vallombrosiens dont le Père Bruno Tozzi, un grand botaniste et membre de la Royal Society de Londres de l’époque, en ce qui concerne la mycologie en particulier, Micheli est considéré à juste titre comme le père de la mycologie moderne.
C’est à lui qu’il faut attribuer le mérite d’avoir découvert et prouvé que même les champignons se reproduisent par le biais des germes avec sa découverte des spores et de leur germination, basée sur la ferme conviction qu’aucune plante ne pouvait se reproduire sans grains et qu’il n’y a pas de plante sans fleur et sans graine. Micheli a découvert les cheilocystides et les pleurocystides à l’aide d’un microscope rudimentaire à trois objectifs qui lui imposait de changer d’objectif pour en augmenter l’agrandissement. Micheli nous décrit ce qu’il vit comme suit:
“…je pouvais voir que de minuscules grains étaient distribués de manière équidistante les uns par rapport aux autres; ce qui me frappait le plus c’était que chacun d’eux était placé sur une base me faisant dire, non sans doutes d’ailleurs, qu’ils pouvaient être les fleurs ou les calices des champignons. ”
De plus:
“…dans le seuil de la lame…naissent des fleurs sans pétales, nues, formées par un simple filament cylindrique; ces fleurs sont solitaires dans certaines espèces, c’est à dire désunies, alors qu’elles sont regroupées pour former des masses ou des rubans dans d’autres. Des grains ronds ou circulaires, de dimensions infimes, naissent un peu partout sur la surface de la lame. Dispersés singulièrement, chez certaines variétés, ils sont groupés quatre par quatre chez d’autres. ”
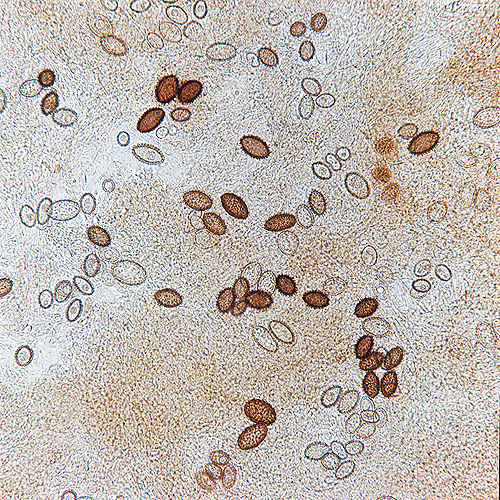
Spores de Tuber melanosporum observées au microscope © Giuseppe Mazza
“…certaines variétés de champignons, plus particulièrement celles qui naissent de déjections équines, bovines ou autres, nous ont fait remarquer quelque-chose qui mérite considération: le fait que la face des lames ne comporte pas uniquement des grains mais également des corps diaphanes, de forme conique ou bien pyramidale, qui paraissent avoir été conçus par la nature pour empêcher que les lames se touchent entre elles en retenant ainsi les grains murs et en les empêchant de tomber…”.
L’histoire de cette matière fascinante s’enrichit considérablement au XIXe et XXe siècles, grâce aussi à la découverte de nouvelles méthodes et de nouveaux instruments. Cependant, nous allons nous arrêter là car il serait trop long de raconter toute l’histoire de la mycologie après Micheli. En effet, nous avons connu de grands botanistes systématistes et mycologues, de LINNÆUS à PERSOON en passant par FRIES et bien d’autres encore jusqu’à arriver à L’ABBÉ GIACOMO BRESADOLA qui a été le mycologue le plus important de la fin du XIXe et du début du XXe. Il a étudié les champignons du monde entier, de par les relations qu’il entretenait avec les plus grands savants de la planète, sans avoir jamais quitté Trente!
Pourtant, il faut quand même donner quelques précisions supplémentaires au sujet du grand Carl von Linné, né le 23 mai 1701 à Rashult, dans la province de Småland, en Suède, et mort à Uppsala en janvier 1778, mieux connu sous le nom de Linnæus.
Dans son œuvre principale, “Systema naturae” (1735), il illustre pour la première fois sa méthode de classification taxonomique des royaumes : animal, végétal et minéral. Son œuvre “Species Plantarum” de 1753 introduit la nomenclature binominale où, chaque animal, chaque plante et chaque minéral sont indiqués par deux noms latins : le premier pour qualifier son genre et le second pour l’espèce. La révolution provoquée par le système de Linnæus est la cause de l’organisation du premier Congrès International de Botanique à Paris en 1867 au cours duquel on adopta les “Lois de la nomenclature botanique”.
Le deuxième Congrès International de Botanique qui eut lieu à Vienne en 1905, vit l’adoption des “Règles Internationales de Nomenclature Botanique” (le premier code). On a organisé depuis, tous les 5 ans, des Congrès de niveau mondial pour introduire d’éventuelles modifications du code ; le dernier d’entre eux a eu lieu à Vienne en 2005. Enfin, l’année 2006 a vu la publication du dernier Code International de Nomenclature Botanique qu’on appelle Vienna Code.
Méthodes de nutrition des champignons © Maria Tullii
LE CHAMPIGNON
Le champignon est un organisme vivant formé de cellules unies entre elles et communicantes, pour former une structure constante, diversifiées entre elles en fonction de leur rôle biologique. L’organisme du champignon proprement dit est constitué d’un ensemble de filaments (qu’on appelle des hyphes) qui se répandent dans le substrat de culture, souvent sur plusieurs mètres. L’ensemble des hyphes est appelé “mycélium”.
Les hyphes ne sont pas visibles à l’œil nu individuellement, toutefois, ils s’entrelacent pour former des agglomérats mycéliens : “rhizomorphes” ou “sclérotes”. Les hyphes s’allongent dans la terre par le biais de la subdivision de la cellule apicale ou des cellules intermédiaires qui sont à l’origine des ramifications latérales. Le mycélium peut, de cette manière, accroître sa surface et sa capacité d’absorption des substances nutritives. Ce qu’on appelle couramment champignon, celui qu’on récolte, n’est autre que le fruit produit par le mycélium lorsque les conditions d’humidité et de température sont favorables. Le fruit est plus précisément appelé “carpophore” ou “sporophore” et il est aussi constitué par les agglomérats de hyphes. Comme dans le cas de tous les autres champignons, sa fonction est celle de la reproduction que nous allons examiner en détail plus loin.
Cycle de reproduction des champignons © Maria Tullii
L’HABITAT
Les champignons poussent dans presque tous les habitats. Ils peuvent privilégier les endroits ouverts comme les environnements très humides, les terrains acides ou bien basiques, la coexistence avec les essences arborées ou arbustives ou encore les prés ; bref, quelle que soit la nature d’un habitat on trouvera une ou plusieurs espèces de champignons aptes à le coloniser.
De plus, il y a des espèces de champignons qu’on nomme “ubiquitaires” et qui peuvent coloniser des habitats divers. Quant à l’habitat, il influe sur l’aspect et la croissance des carpophores : une même espèce peut avoir des dimensions et des aspects morphologiques différents en fonction de l’altitude, du degré d’humidité et de l’exposition au soleil.
LES CONDITIONS DE CROISSANCE
La croissance de l’organisme du champignon est très dépendante des conditions ambiantes : lorsque l’humidité et la température sont favorables, le mycélium produit des carpophores qui, en se développant, produisent des spores qui, donneront vie à un nouveau cycle à leur tour, en tombant au sol. Les conditions de croissance varient de zone à zone : la période de croissance des champignons sera d’autant plus brève que le climat est rude et la saison hivernale est longue alors qu’elle sera quasiment nulle dans les régions sèches en permanence et quasiment perpétuelle dans les régions tempérées.
LA REPRODUCTION
Comme nous le disions, le mycélium produit des fruits et donne naissance à des carpophores en présence de conditions climatiques d’humidité et de température idéales. Pour se reproduire, ceux-ci, par le biais de cellules spécialisées, créent des millions de grains qu’on appelle des “spores” qui, une fois dispersées dans l’environnement deviennent l’origine d’un nouveau cycle de vie (même à très grande distance) avec l’aide de facteurs extérieurs tels que le vent, la pluie ou les vecteurs animaux. Les spores sont des cellules minuscules, non visibles à l’œil nu et qu’on ne peut donc observer qu’au microscope. L’unité de mesure des spores est le micron (µm) qui correspond à un millième de millimètre.
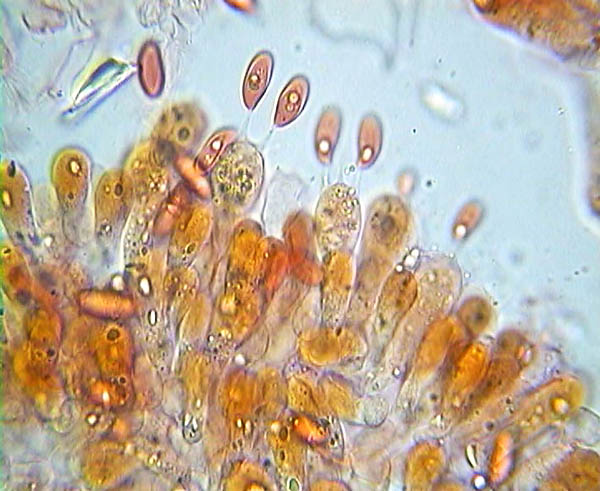
Basides et spores de Lepiota alba © Pierluigi Angeli
Les spores sont sexuées (au moins chez les champignons supérieurs que nous sommes en train d’examiner), non pas dans le sens traditionnel de mâle ou femelle, mais dans le sens de la charge différente qu’ils possèdent : positive (+) ou négative (-).
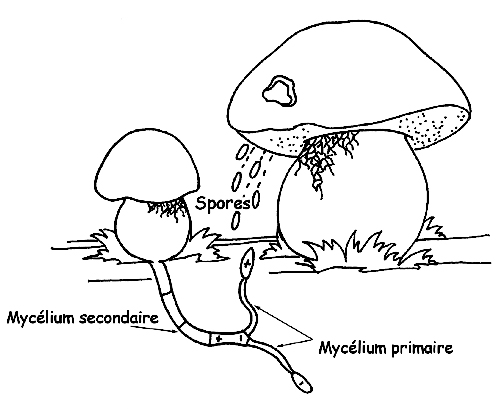
Lorsqu’une spore trouve un substrat adapté à sa reproduction en condition favorable, elle pousse en donnant naissance à un filament qui a la même charge que la spore d’origine (hyphe) qui peut se développer et pousser (mycélium primaire) mais ne peut donner de fruits. Pour que le mycélium primaire puisse fructifier il doit pouvoir se joindre à un autre mycélium primaire de la même espèce mais de charge opposée.
L’union des deux mycéliums, positif et négatif, s’appelle mycélium secondaire qui constitue l’organisme de “champignon” proprement dit.
Cependant, comme la possibilité qu’un mycélium primaire donne lieu à une nouvelle colonie de champignons est très petite, chacun des corps producteurs de fruits émet des millions de spores.
LES SYSTÈMES DE NUTRITION
Toutes les formes de vie qui existent sur terre se divisent en PRODUCTEURS ou AUTOTROPHES : organismes vivants qui sont en mesure de se nourrir de manière autonome en synthétisant les substances dont ils nécessitent, à partir d’éléments organiques présents dans la nature. Les plantes se servent de la lumière pour produire des matières complexes (comme les sucres) en partant de substances simples comme le dioxyde de carbone et l’eau, par l’intermédiaire du processus de Photosynthèse chlorophyllien. CONSOMMATEURS ou HÉTÉROTROPHES : organismes vivants qui se procurent les substances nécessaires à leur survie à partir de producteurs ou autres consommateurs.
Les sources qui sont l’origine de leur nourriture nous permettent de classer les consommateurs en quatre groupes : Herbivores : se nourrissent de substances végétales ; Carnivores : se nourrissent des autres consommateurs ; Omnivores : se nourrissent aussi bien des autres consommateurs que de substances végétales ; Décomposeurs : se nourrissent à partir d’organismes, morts ou vivants, en transformant les matières organiques complexes en substances simples comme le dioxyde de carbone et l’eau.

Clitocybe odora, un champignon décomposeur de litière © Giuseppe Mazza
Les plantes supérieures, celles qui sont en mesure de s’autoalimenter en transformant les substances organiques (l’eau et le dioxyde de carbone) en substances complexes à travers le processus de photosynthèse chlorophyllienne, font partie du groupe des organismes Autotrophes.
Les champignons, autrefois classés parmi les végétaux, font désormais partie d’un domaine qui leur est propre : le Regno Fungi, puisqu’ils n’ont pas la capacité d’assurer la fonction chlorophyllienne. Il faut donc les affecter au groupe des organismes Hétérotrophes et à la subdivision des Décomposeurs.
Sur la base du système nutritionnel adopté, les champignons se classent en :
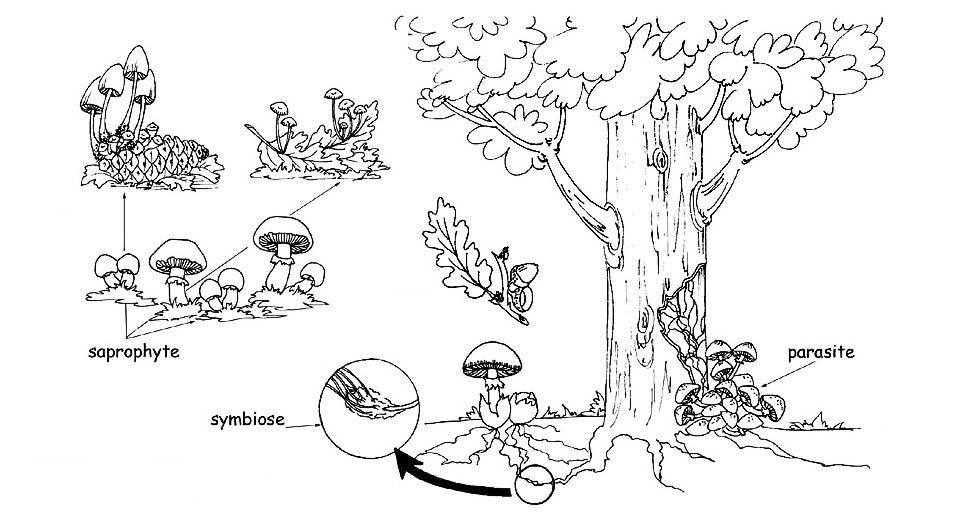
►Saprophytes : se nourrissent de matières organiques en décomposition, aussi bien d’origine végétale qu’ animale. Ils effectuent un premier cycle de désagrégation et préparent le terrain pour les bactéries qui compléteront ce processus de manière à ce que tous les résidus organiques soient transformés en substances inorganiques : l’eau, le dioxyde de carbone et les sels minéraux. Les champignons qui naissent sur les prés, les pâturages, les feuilles, les branches sèches et les organismes des animaux morts, font partie de ce groupe. Nous pouvons considérer les champignons saprophytes comme les opérateurs écologiques de la nature : sans eux, les détritus végétaux et animaux suffoqueraient l’environnement.
►Parasites : vivent aux dépens d’autres êtres vivants, aussi bien végétaux qu’animaux. Si d’un côté les champignons parasites sont une véritable menace pour les plantes et les animaux car ils provoquent des pathologies graves en mesure d’amener la mort de ceux-ci, d’un autre côté ils agissent comme éléments régulateurs en supprimant les plantes les plus faibles ou malades pour favoriser le développement de celles qui sont saines. Une fois que l’organisme hôte, qu’il soit animal ou végétal, est décédé le champignon parasite se transforme en saprophyte pour achever la démolition de la substance organique morte.
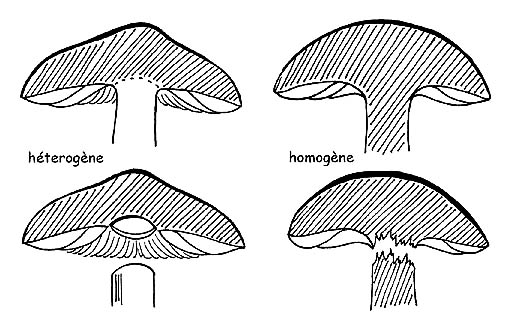
Homogénéité du champignon © Maria Tullii
►Myrorhiziques ou symbiontes : sont les champignons qui vivent en symbiose avec les plantes supérieures en instaurant avec elles un rapport qui est mutuellement bénéfique. Les parties apicales du mycélium enveloppent la zone terminale des poils radicaux des plantes supérieures en augmentant leur surface d’absorption. La plante dispose ainsi d’une quantité supérieure de matières inorganiques qu’elle transformera, grâce à la photosynthèse chlorophyllienne, en substances organiques à la disposition du champignon. De plus, la “mycorhize” (formée par les poils radicaux qui sont enveloppés par les hyphes fongiques) protège les racines de la plante des attaques de pathogènes en les renforçant et en les rendant plus saines. Les mycorhizes peuvent être : Endomycorhizes, lorsque les hyphes fongiques pénètrent les espaces intercellulaires et l’intérieur des cellules végétales. L’expansion du champignon s’effectue à travers les cellules corticales bien qu’elle ne morde pas sur le cylindre central ni sur les cellules apicales de la racine. Ectomycorhyzes, quand les hyphes fongiques encerclent la racine en pénétrant les espaces intercellulaires mais non pas l’intérieur des cellules végétales. Le tressage des hyphes, autour des cellules radicales, est connu sous le nom de réticule de Hartig.
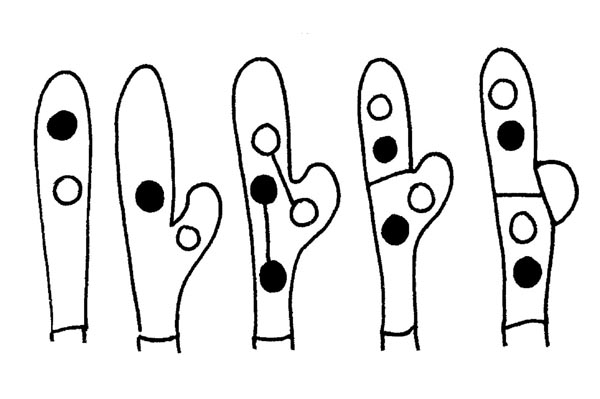
MORPHOLOGIE DES CHAMPIGNONS
Pour qu’on puisse attribuer à un champignon un nom et un prénom corrects (Genre, espèce), il faut d’abord en identifier toutes les caractéristiques. Commençons par distinguer certains types des champignons : Agaricomycètes : sont tous les champignons composés d’un chapeau et d’une tige ; Ascomycètes : ils ont soit une pseudo-tige ou un pseudo-chapeau ou bien ils ont la forme d’une coupe, ils peuvent être sessiles ou non-sessiles ou encore ils peuvent avoir la forme d’un tubercule dans quel cas ils sont, hypogés ou semi-hypogés, Gastéromycètes : de forme ± ronde avec ou sans pseudo-tige, hypogés ou épigénétiques.
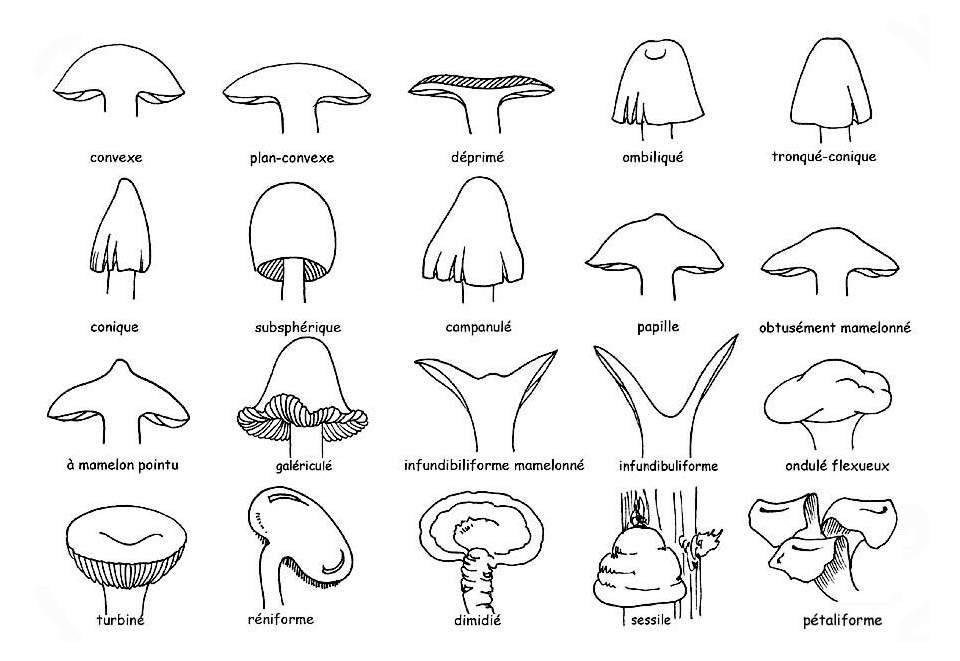
Forme du chapeau © Maria Tullii
Faisons maintenant quelques considérations au sujet des spécificités morphologiques des champignons qui sont munis d’un chapeau et d’une tige : les différences sont parfois minimes et seule une analyse minutieuse permet une identification exacte.
HOMOGÉNÉITÉ
Le carpophore est formé par le tressage des hyphes qui sont unis étroitement ou au moins disposés parallèlement entre eux. Nous sommes en présence d’un champignon homogène si la structure du chapeau et de la tige est la même. Cela sera confirmé par une chair qui s’effrite lorsqu’on essaye de séparer la tige du chapeau. Par contre, si les hyphes du chapeau et de la tige sont structurés de manière différente, il sera possible de désunir ces deux parties sans ruptures, comme si elles étaient juste appuyées l’une sur l’autre. On dira alors que le champignon est hétérogène.

Marge et profil du chapeau © Maria Tullii
CHAPEAU
On observera du chapeau :
► ses dimensions, c’est le diamètre atteint à la fin de la croissance ;
► sa forme, qui varie beaucoup entre la jeunesse et la maturité.
Normalement ce sont les spécimens adultes que l’on considère bien qu’on ferait mieux d’observer plusieurs spécimens à différentes étapes de croissance. Habituellement, chez les exemplaires jeunes, le chapeau est plus renfermé pour ensuite s’ouvrir et s’aplanir jusqu’à ce qu’il atteigne sa forme typique de l’âge adulte ;
► la marge, qui est toujours importante et même déterminante chez certaines espèces. Elle doit être observée scrupuleusement par rapport au plan horizontal du chapeau. Elle peut être droite et donc sur le même plan ou enroulée, tournée vers l’intérieur ; réfléchie, tournée vers l’extérieur ; incurvée, tournée vers le bas ; la marge peut avoir un profil : régulier, ondulé, lobé, festonné, sinueux ou plissé, entier ou strié ; puis, il est considéré excédentaire lorsqu’il dépasse, même légèrement, l’hyménium (lames, tubes, etc.) ;
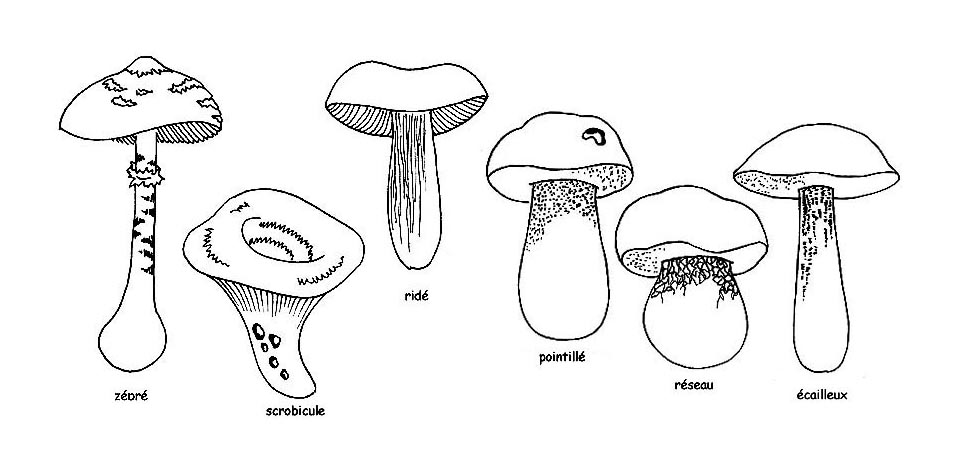
► la cuticule du chapeau, peut être : lisse, poisseuse ou visqueuse, tomenteuse, velue, veloutée ou rugueuse. On remarquera si elle peut être séparée de la chair du chapeau : la cuticule qui ne se détache pas est dite adnée ou non séparable alors que si elle se détache entièrement elle est séparable. Par contre, si elle ne se sépare qu’en partie on dira qu’elle est partiellement séparable et on se concentrera sur la portion du rayon du chapeau qui est découverte (1/ 2, 1/3, etc.)

Décorations du chapeau © Maria Tullii
► la surface du chapeau peut avoir, ou pas, des décorations. Une surface qui ne comporte pas de décorations peut être unie, lisse, uniforme alors que si elle en comporte celles-ci pourront être :
Innées, soit, elles sont une partie intégrante de la structure de la pellicule comme les fibrilles, les vergetures ou les tâches ; Adnées ou appliquées, c’est-à-dire qu’elles sont dérivées de la dissociation de la pellicule comme les fêlures et les écailles, ou bien des résidus du voile général comme les verrues, les plaques, la poudreuse, la pilosité, la pruinosité ;
► la couleur est une caractéristique importante mais non pas capitale dans la mesure où bon nombre de champignons passent par des variations chromatiques importantes en fonction des conditions climatiques, de leur âge et de leur exposition. Pour décrire les couleurs, nous allons nos référer à celles que nous connaissons habituellement et que nous associons à nos objets quotidiens (rouge brique, vert olive, etc.).
HYMÉNOPHORE
L’hyménophore est l’ensemble des structures macroscopies et microscopiques préposées à la reproduction des spores.
Il est d’une importance fondamentale non seulement du point de vue biologique mais aussi pour celui de la détermination de l’espèce. Chez les champignons à tige et chapeau, il est situé, en général, dans la partie inférieure du chapeau. Il peut être formé par des lames, des tubes, des pores et des aiguillons.
Accumulation des spores © Maria Tullii
Chez certaines variétés, il est presque lisse ou à peine rugueux.
Alors que l’hyménium est exposé à l’air dans la majeure partie des cas, chez les champignons qui appartiennent à la famille des Gastromycetidae, par contre, il est protégé et renfermé à l’intérieur du carpophore (les spores seront libérées par la rupture du péridium à la maturation) tout comme pour les Tuberales, chez les ascomycètes.
►Les lames : la première caractéristique que nous devrons observer est la couleur des lames qui peut être : innée, c’est-à-dire la couleur qui est visible est celle de la lame ; dérivée, lorsque la couleur des lames est celle des spores.
Leur couleur une autre caractéristique très importante car elle détermine la désignation d’un champignon à un groupe génétique ou à un autre en fonction de son appartenance à un groupe de chromosporés ou un autre.
Détail de la lame © Maria Tullii
Les groupes des chromosporés sont cinq : leucosporés, c’est-à-dire à sporée blanche ou blanchâtre ; à sporée ocre ; rhodosporés, à sporée rose ; à sporée violette et mélanosporés, à sporée noire.
La couleur de la sporée est vérifiable par la récolte de la poudre sporale sur un support de couleur contrastante.
Il faudra observer ensuite : l’ homogénéité des lames : comme pour le chapeau et la tige il faudra remarquer si les lames se détachent facilement de la chair du chapeau : dans le cas affirmatif on parlera de lames hétérogènes ou séparables ; par contre si elles ne se détachent pas mais elles se cassent au cours de cette tentative, on dira qu’elles sont homogènes ou bien non séparables.
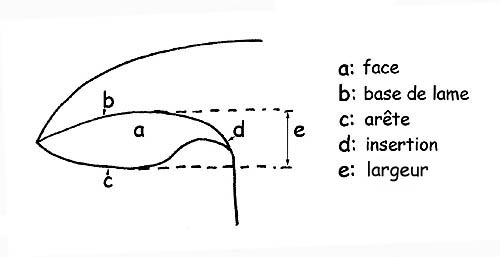
La forme des lames en fonction de ses composantes : bord, arête, largeur, épaisseur, etc., et l’ insertion de la tige est comme suit :

Insertion de la lame au pied © Maria Tullii
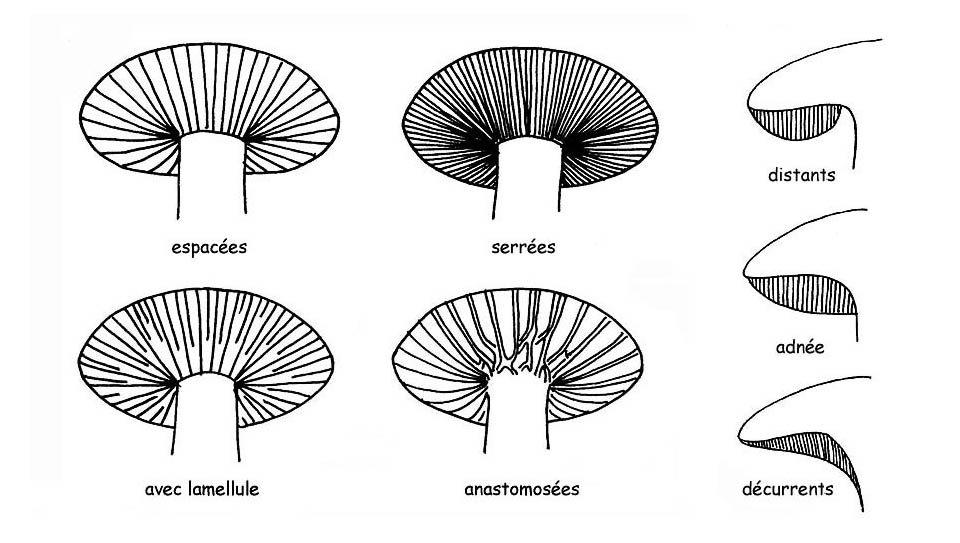
►libres ou distantes, lorsque les lames n’atteignent pas le stipe, une caractéristique commune à tous les champignons hétérogènes.
►adnées, quand les lames sont attachées au pied sur toute leur largeur.
►échancrées ou crochetées, lorsqu’elles ne sont attachées au stipe que sur une petite portion de leur largeur.
►décurrentes, lorsqu’elles sont attachées au stipe sur une portion qui est supérieure à leur largeur.
►décurrentes par une dent, quand une partie de la lame n’est décurrente que par une petite prolongation (dent).
►collarium, lorsque les lames, éloignées du pied, lui sont reliées par un petit cordon circulaire.

Profil de la lame © Maria Tullii
En observant le profil de la lame depuis son arête on peut définir la lame comme : aigue, sinueuse, ventrue, obtuse ou droite.
La base de la lame est l’arête de la lame qui est attachée à la chair du chapeau.
L’arête de la lame (c’est l’arête qui se trouve à l’opposé de la base de la lame) peut être entière, échancrée, dentelée, lisse, floconneuse ou pruineuse. Elle peut être de la même couleur que les lames ou bien de couleur différente en quel cas c’est une arête hétéromorphe.
Par contre, si l’arête de la lame se divise à proximité du stipe, ce sera une lame fourchue avec présence possible de Lamelles : ce sont des lames qui, bien que partant de la marge du chapeau, n’atteignent pas le pied.
Elles s’alternent souvent avec les lames et sont de longueur variable.
La partie terminale des lamelles peut être tronconique, arrondie ou sinuée ; il peut y avoir Anastomose : les lamelles sont reliées par des joints transversaux qui unissent les lamelles entre elles à plusieurs endroits.
Pour plus de clarté : on entend par largeur la distance qu’il y a entre la base et l’arête de la lame.
Typologie des lames et des tubes © Maria Tullii
Par épais ou fin on qualifie l’épaisseur de la lame. Par espacées ou serrées on se rapporte à la distance qui sépare les lames. On qualifiera de la même manière l’hyménium sauf pour les modifications qui s’imposent aux tubes et pores.
On observera la couleur des tubes et des pores de spécimens jeunes et mûrs (les tubes et les pores peuvent être aussi bien de la même couleur que de couleur différente), avec toutes les variations intermédiaires : la longueur des tubes, la manière dont ils sont attachés au stipe, la grandeur des pores, leur forme, etc.
Insertion du stipe © Maria Tullii
Remarque : chez les hyménophores et les tubes avec pores, l’ homogénéité est la caractéristique qui détermine leur appartenance à un groupe : chez les Boletales la partie hyménale est hétérogène et donc facilement détachable ; chez les Poliporales la parti hyménale est bien attachée à la chair et donc elle ne peut en être séparée.
STIPE
Si le carpophore est composé d’un pied et d’un chapeau on dira qu’il est pédonculé, par contre, s’il n’a pas de stipe on le qualifiera de sessile.
Il y a de nombreuses spécificités qu’il faut observer aussi pour le stipe.
Elles jouent toutes un rôle fondamental dans la détermination de l’espèce.
On observera :
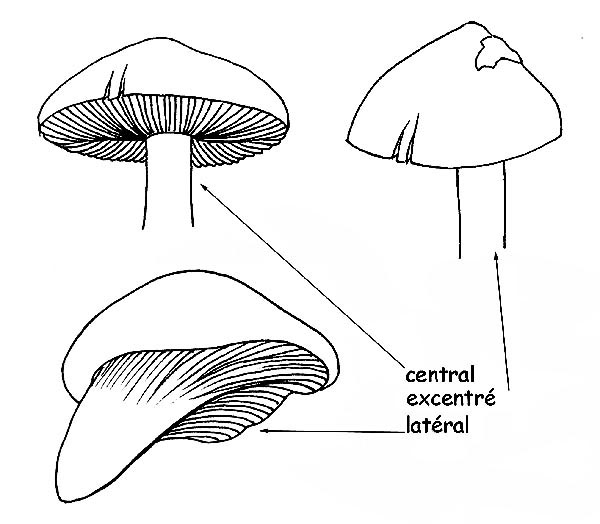
►l’insertion au niveau du chapeau qui peut être centrale, latérale ou même excentrée.
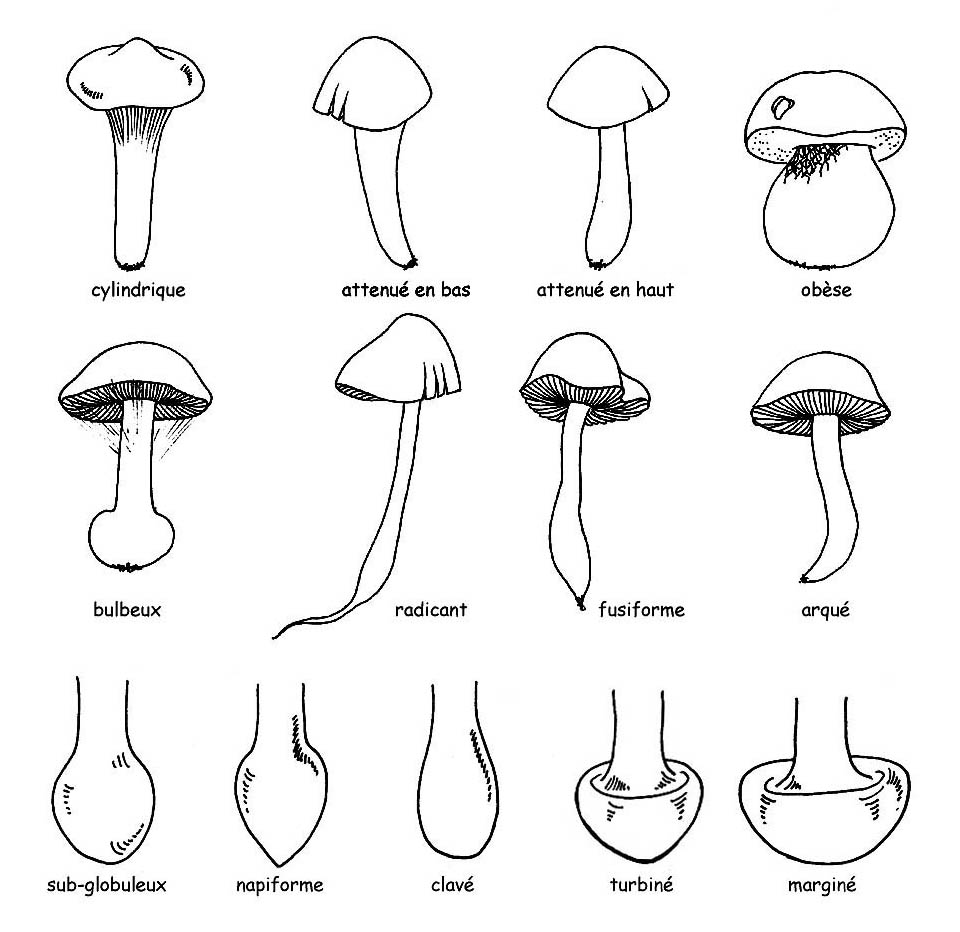
►sa forme : cylindrique, atténuée en bas ou en haut, obèse, ventrue ou bulbeuse
►le bulbe à la base peut être défini, en fonction de sa forme : sub-globuleux, napiforme, clavé, turbiné ou marginé
►son intérieur peut être soit, pulpeux, fibreux-pulpeux, fibreux, plein ou creux, fistuleux, cortiqué, etc.
►Il peut encore y avoir des décorations de type différent. La surface du pied peut être lisse, floconneuse, squameuse, fibrilleuse, verruqueuse, velue, pointillée ou clavée ou avec d’autres décorations; mais aussi sèche, visqueuse ou glutineuse.
Forme du stipe et typologie du pied © Maria Tullii
Toutefois, il faudra faire des considérations à part pour les Boletaceae dont les pieds arborent des maillages.
On devra dans ce cas observer : la couleur, la grandeur des mailles, si elles sont ± allongées ou si elles sont ± en relief.
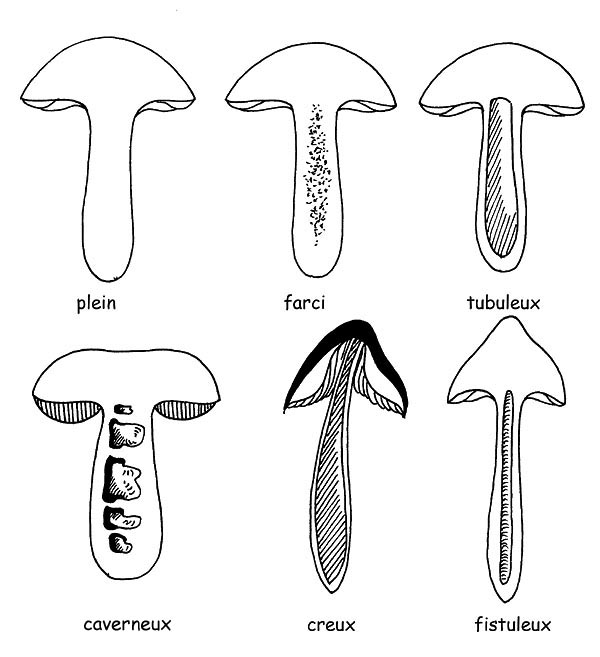
Intérieur du pied © Maria Tullii
Le maillage peut être totalement absent ou bien le pied peut être recouvert de pointillages ou de fibrilles ou encore de rugosités ou d’ écailles comme dans le Leccinum.
VOILES
Tous les carpophores, dans le premier stade de leur vie (primordium) sont enveloppés par une membrane fermée (voile général).
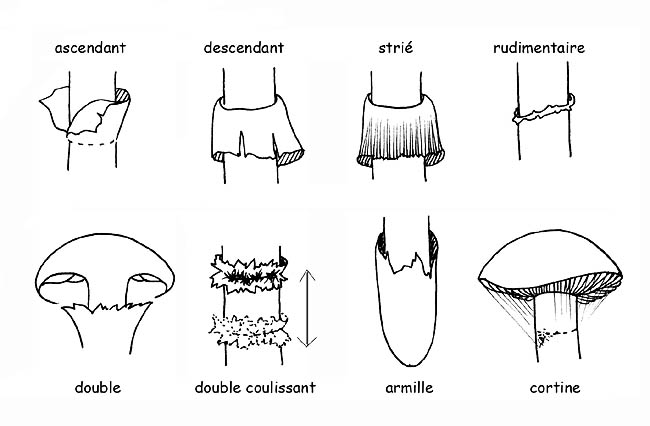
Chez la plupart des champignons ce voile disparaît dès sa première phase de développement mais, pour d’autres espèces qui appartiennent au genre des Agaricaceae, le voile général persiste jusqu’à l’âge adulte et il reste visible sur le carpophore sous différentes formes : des verrues ou des plaques sur le chapeau, la volve ou l’ armille à la base du pied ; certaines variétés d’ Agaricaceae ont un autre voile, le voile partiel, qui recouvre la partie inférieure du chapeau en partant de la marge du chapeau et en arrivant jusqu’ au stipe à protection de l’hyménophore.
Ce voile, d’abord fermé sue les exemplaires jeunes, se détache de la marge du chapeau au fur et à mesure que celui-ci s’ouvre en restant attaché au pied où il reste ± apparent sous forme d’ anneau.
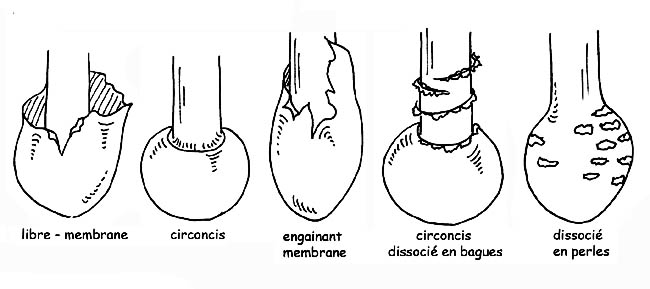
La volve peut être : en forme de sac, membraneuse, engainante, circoncise, dissociée en bagues ou perlée ; l’anneau peut être floconneux, large, étroit, infère (lorsqu’il se détache de bas en haut), supère (lorsqu’il se détache de haut en bas), etc. L’armille est le résidu du voile.
Certaines espèces arborent une double bague, résidus du voile général et du voile partiel qui sont unis l’un à l’autre au niveau du stipe ; chez d’autres types, le résidu du voile partiel est visible par des filaments très fins et fugaces (cortine).
Décorations du pied © Maria Tullii
CHAIR
La pulpe d’un champignon, c’est-à-dire cette partie qu’on voit lorsqu’on coupe un champignon de haut en bas s’appelle la chair. Comme pour les autres parties du champignon, la chair aussi possède des particularités fondamentales pour la détermination de son espèce : sa consistance peut être : ferme, compacte, molle, coriace, membraneuse, cireuse, etc. sa couleur et éventuellement son virage (mutation de la couleur de la chair exposée à l’air due à l’oxydation de deux enzymes), une particularité qu’il faut observer avec attention en prenant note de son intensité et le temps d’apparition du virage (très important dans les Boletaceae). Il peut avoir une tonalité bleue ou rouge en fonction de la domination d’un enzyme sur un autre.
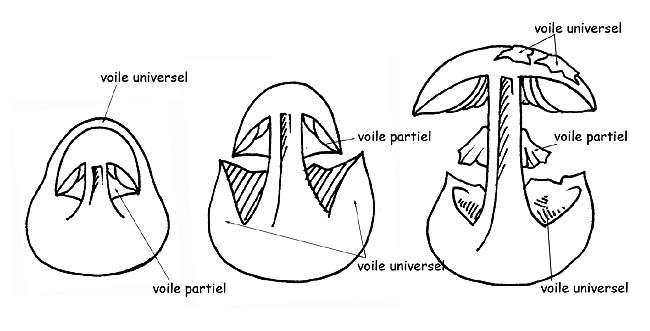
Développement des voiles © Maria Tullii
Le degré du virage sera plus fort chez les exemplaires jeunes et frais et plus faible ou inexistant pour les exemplaires plus vieux ; l’ hygrophanéité : la capacité ± apparente de la chair à absorber l’eau ; la reviviscence : la capacité de la chair de sécher et de réacquérir sa fraicheur au contact de l’eau ; les propriétés organoleptiques : le goût et l’odeur de la chair sont des qualités importantes et même capitales chez certaines espèces du type Agaricus .
GÉNÉRALITÉS
SUR LA SYSTÉMATIQUE
La mycologie est une branche des Sciences Naturelles qui traite les champignons. Comme toutes les sciences, elle doit se servir de certains instruments de recherche et de catalogage pour être claire. Les voici :
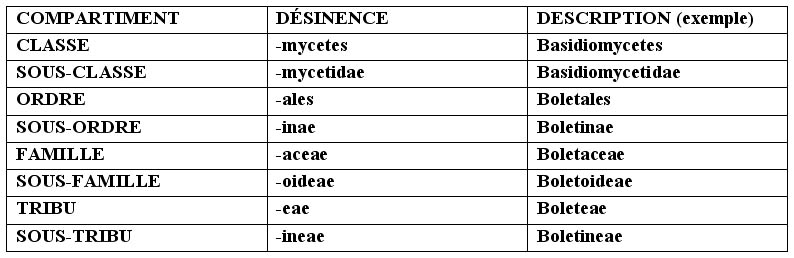
► la SYSTÉMATIQUE : c’est l’instrument scientifique qui permet de placer le sujet d’une étude au centre d’un plan particulier (système) qui est caractérisé par une subdivision en compartiments, chacun avec des qualités communes, à l’intérieur desquels on trouve d’autres compartiments et ainsi de suite jusqu’à ce qu’on arrive au niveau de l’unité (c’est une sorte de système de boîtes chinoise où l’espèce se situe à l’intérieur d’un genre, le genre à l’intérieur d’une famille, etc. voir tableau au bas de la page). C’est un instrument en phase de mise à jour constante et qui n’est pas univoque dans la mesure où il peut être interprété de manière différente par les mycologues. Le classement que nous proposons est celui des pères de la systématique moderne.
Typologie de la volve © Maria Tullii
► la TAXONOMIE est la science qui étudie la classification du sujet étudié soit, le positionnement correct des différents taxa au sein du plan systématique. On entend par taxon le regroupement de tout degré (espèce, genre, famille, ordre, classe, division, règne).
► la NOMENCLATURE, est l’ensemble des règles qui permet l’attribution d’un nom scientifiquement valable et correct au sujet étudié. Ces règles sont sanctionnées par le Code International de Nomenclature Botanique (C.I.d.N.B). C’est à ce code que tous les savants et les auteurs de livres et de publications diverses sur la botanique et la mycologie se réfèrent. Le C.I.d.N.B. a été fondé en 1867 ; des Congrès et des Comités de niveau mondial sont organisés tous les 5-6 ans pour introduire des mises à jour et des modifications éventuelles.
CLASSEMENT ET PLAN SYSTÉMATIQUE DES CHAMPIGNONS
Nous proposons ci-dessous un plan systématique pour le classement des champignons en nous référant aux écoles de R. Kühner et de R. Singer, qui sont des chercheurs de mycologie célèbres, et aux règles du C.I.d.N.B (Code International de Nomenclature Botanique).
Typologie de l’anneau © Maria Tullii
Remarque : lorsqu’on cite une espèce, le nom de cette dernière doit être écrite dans sa forme binominale latine ; le premier nom, celui du Genre doit comporter une initiale majuscule (ex. Agaricus), le second, celui de l’espèce doit être écrit en minuscule (ex. campestris) ; ensuite c’est généralement le nom de l’auteur, en abrégé, qui a décrit et classé le champignon (ex. Bull. pour Bulliard) ; si (à l’intérieur d’une parenthèse), le nom est suivi d’un autre nom, il peut s’agir de celui du sanctionnateur qui dans ce cas devra toujours être précédé de “ :”et il ne peut être que Persoon ou Fries, (es. Bull. :Fr.), ou bien de la personne qui s’est occupée du diagnostic original, dans le cas d’absence de l’auteur (par exemple, l’absence du diagnostic latin, et dans ce cas le nom est précédé par “ex” (ex. Bull. ex Vent.).
Il peut y avoir aussi un troisième nom (hors parenthèses), qui représente l’auteur qui a modifié le positionnement du Genre ou de l’espèce (ex. Bull. : Fr.) Kumm. La désinence identifie les différents compartiments de la systématique. Le Genre, le sous-genre, la section, l’espèce, la variété et la forme ont des désinences variables, liées à la racine linguistique. Le Règne et la Division dérogent à cette règle puisqu’elles n’on pas de désinence particulière. En partant de la Classe pour aller à la Sous-tribu nous avons en ordre décroissant :
MICROSCOPIE
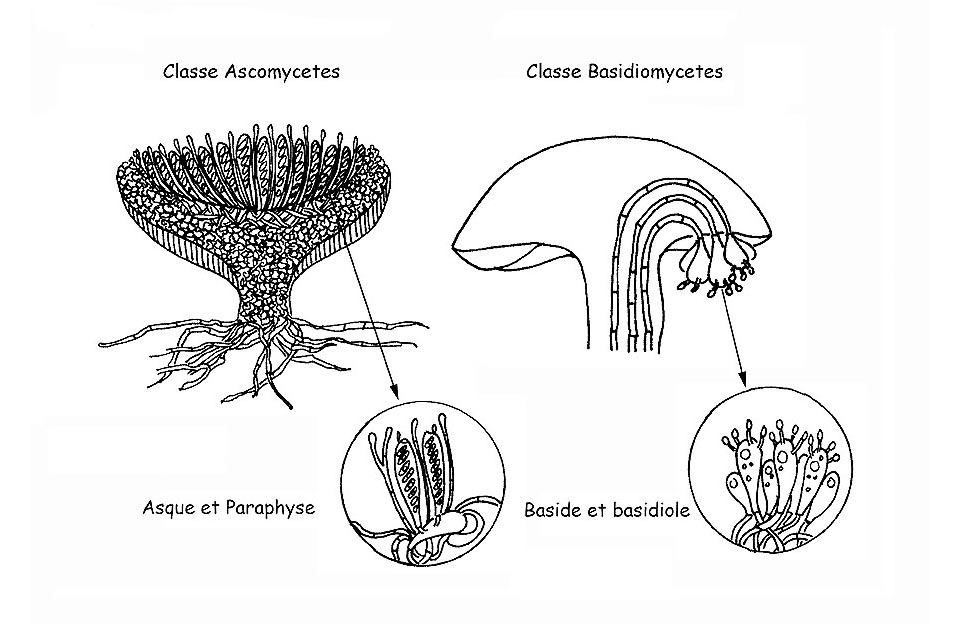
Les propriétés microscopiques des champignons sont tout aussi importantes que celles macroscopiques pour une identification précise de l’espèce en question. Chez certains genres, la microscopie est le seul instrument sûr qui permette l’identification de leurs nombreuses espèces (ex. Le Genre Mycena). Cet exercice se veut être davantage une dissertation descriptive qu’un véritable traité scientifique au cours de laquelle nous allons illustrer les structures microscopiques principales que l’on trouve dans les champignons supérieurs qui sont partagés en deux grandes classes distinctes : Ascomycetes et Basidiomycetes . Nous allons aussi décrire également les réactifs et les techniques les plus fréquentes utilisées pour leur observation.
Champignons supérieurs © Maria Tullii
ASCOMYCÈTES: LEURS ÉLÉMENTS MICROSCOPIQUES
Ascomycètes © Maria Tullii
Il est important, pour cette classe de carpophores, d’au moins évoquer le développement de la partie fertile du champignon : l’hyménium (thèce).
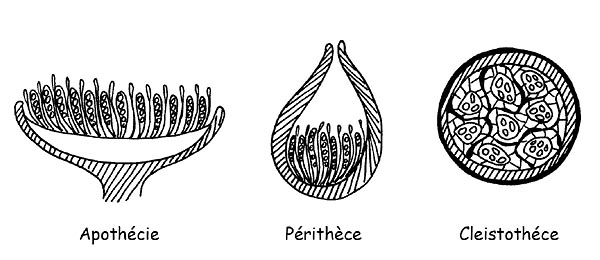
Lorsque le carpophore expose complètement à l’air libre sa partie hyménale, on se alors trouve en présence d’un Apothèce alors qu’on parlera de Périthèce quand l’hyménium ne communique avec l’extérieur qu’à travers une petite ouverture apicale.
Enfin, lorsque la partie hyménale ne communique pas du tout avec l’air ambiant alors aura affaire à un Cleistothèce (dans ce cas la majorité des espèces connaissent un développement hypogé ou semi-hypogé).
Nous allons voir pourquoi ces distinctions sont importantes d’un point de vue microscopique pour une observation particulière des différents éléments.
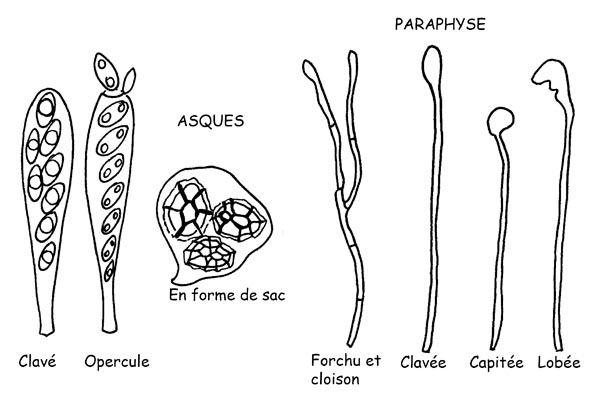
Asques et Paraphyses © Maria Tullii
Les Asques desquels dérive l’appellation de la classe, sont des éléments fertiles qui produisent des spores et se développent à partir d’hyphes spécialisés qu’on appelle des ascogènes.
Ils ont une forme cylindriforme ± clavée ou bien ils sont en forme d’outre contenant les spores (généralement celles-ci sont huit même si leur numéro peut varier pour atteindre trente-deux ou plus) jusqu’à leur maturation.
Ils peuvent être équipés d’une paroi simple ou double (tunique) et peuvent réagir positivement ou négativement au réactif de Melzer.
Dans le cas d’une réaction positive on a affaire à des amyloïdes ou iodés positifs.
Les asques cylindriformes, typiques chez les Apothèces et les Périthèces, s’ouvrent à la maturation des spores
Cette ouverture peut être par lacération ou cassure de la partie apicale ou alors par l’ouverture d’un couvercle minuscule (opercule) qui laisse dépas- ser les spores mûres.
Les asques en forme de sac qui sont typiques des Cleistothèces, par contre, libèrent les spores par la décomposition de leurs parois.
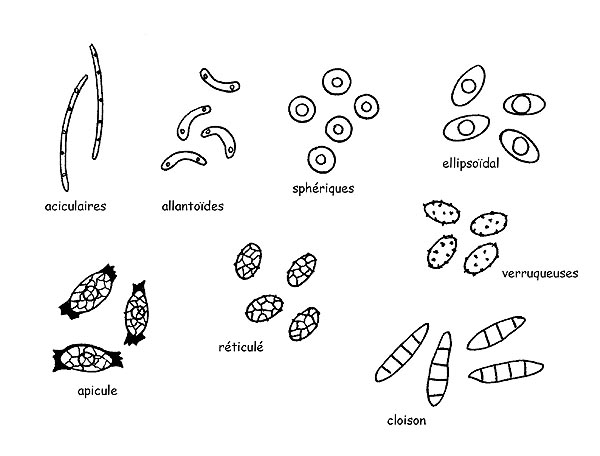
Spores © Maria Tullii
Les Paraphyses sont des hyphes fines et stériles lesquelles, emmêlées aux asques, jouent un rôle de soutien pour ces derniers.
Elles sont principalement cylindriformes mais elles peuvent être aussi fourchues ou positionnées.
Leur sommet peut prendre plusieurs formes et peut être : clavé, lancéolé, capitulé, lobé, incrusté, etc.
On observe la positivité au réactif de Melzer même chez certaines espèces de paraphyses.
Les Spores des ascomycètes peuvent avoir des formes (aciculaire, allantoïde, sphérique, ellipsoïdale, apiculée, etc.) des couleurs et des ornements (réticules, crêtes, verrues, épis, etc.) très variées et elles peuvent être positionnées aussi bien longitudinalement que transversalement.
Les Poils sont un élément de distinction pour bien de variétés d’ascomycètes. Leur rôle est celui de protéger l’hyménophore et de retenir l’humidité nécessaire au développement de l’espèce, ils peuvent être disloqués au niveau de la surface extérieure ou de l’arête du carpophore et peuvent avoir des formes et des dimensions différentes, tout en étant cloisonnés ou non. Leur sommet peut être pointu ou ± arrondi etc.
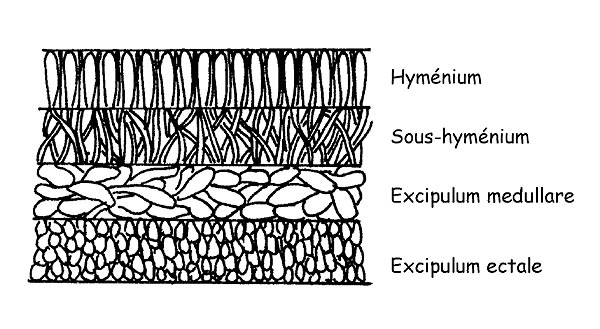
Coupe de l’Apothèce © Maria Tullii
La Texture : on entend par texture la structure de la chair qui peut être composée de cellules de forme et de conformation différentes.
Il est également important d’observer la section complète du carpophore des Apothèces.
Elle est constituée de plusieurs couches: Hyménium, Subhyménium, Excipulum medullare, Excipulum ectale.
L’observation de la configuration des couches simples des hyphes peut être un passage essentiel pour la définition d’une espèce.
BASIDIOMYCÈTES : LEURS ÉLÉMENTS MICROSCOPIQUES
Nous avons vu que les spores des ascomycètes se développent à intérieur de structures spécialisées, les asques, qui les protègent jusqu’à la maturation complète. A l’inverse, les basidiomycètes laissent leurs spores se développer et mûrir à l’extérieur des structures fertiles, les Basides.
La classe des Basidiomycètes se divise en deux sous-classes principales : les Holobasidiomycetes et les Heterobasidiomycetes : Toutes les espèces munies de basides monocellulaires appartiennent à la première espèce alors que la deuxième espèce englobe les espèces munies de basides pluricellulaires.
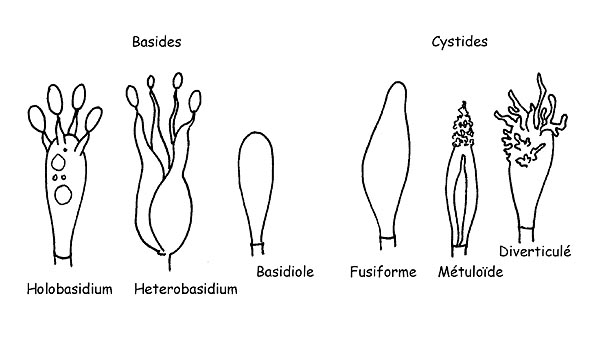
Structures des Basidiomycètes © Maria Tulliii
Les Basides sont des cellules spécialisées dans la production de spores. Elles naissent de la cellule terminale d’un hyphe. L’intérieur de la baside est l’endroit où la division cellulaire a lieu. Celle-ci est l’origine de quatre cellules qui deviendront chacune une spore. La formation des spores a lieu en haut des protubérances qui se trouvent au sommet de la baside (stérigmes). Elles seront lâchées dans l’environnement voisin à la maturité. Les basides sont généralement ± cylindriques et ± clavées, elles comportent généralement quatre stérigmes (basides tétrasporiques) mais elles peuvent en avoir aussi deux ou trois ou plus de quatre ainsi que du pigment cellulaire ou pas.
Les Basidioles ne sont autre que des basides encore en cours de formation.
Les Cystides sont des éléments stériles qui peuvent se trouver aussi bien dans la partie hyménale que dans la cuticule ou sur la stipe.
Les cystides, présentes sur la face des lames, s’appellent des Pleurocystides. Si elles se trouvent sur l’arête des lames elle s’appellent Cheilocystides alors que celles présentes sur la cuticule ou la stipe s’appellent respectivement Dermatocystides et Caulocystides. Les cystides peuvent avoir des formes très différentes, d’espèce à espèce, avec ou sans incrustations, avec une paroi simple ou double et avec ou sans pigment intracellulaire. Elles revêtent donc une grande importance d’un point de vue microscopique dans la mesure où elles permettent souvent une détermination correcte de l’espèce.
Formation d’anses d’anastomoses © Maria Tullii
Les Spores, même dans les basidiomycètes, sont très variables dans leurs formes, leurs couleurs et leurs décorations. Elles comportent généralement une petite protubérance au niveau d’une des extrémités (apicule), qui n’est autre que l’appendice qui ancrait la spore au stérigme, alors que l’autre extrémité peut présenter un pore germinatif, une sorte de petite ouverture par où sortira le nouveau mycélium.
L’amyloïde, soit la positivité au réactif de Melzer et la cyanophilie, c’est-à-dire lorsque la paroi sporale se colorie de bleu au contact du réactif Bleu coton lactique, sont également aussi importantes pour l’identification des spores des basidiomycètes.
Les Anses d’anastomoses sont le résultat d’un type de croissance particulier des hyphes par l’intermédiaire d’un diverticule produit par la cellule apicale. Les anses d’anastomoses ne sont pas présentes pour tous les genres ou pour toutes les espèces d’un genre alors que leur présence ou absence est souvent déterminante pour l’identification de l’espèce observée. Les anses d’anastomoses se situent généralement au niveau des cloisons des hyphes mais aussi à la base des basides ou des basidioles. Chez les basidiomycètes, l’étude de la forme et de la disposition des hyphes, qui constituent le Pileipellis (revêtement du chapeau), a une importance particulière, surtout pour certains genres comme la Russula et la Caulopellis (revêtement du stipe) comme pour le genre Mycena, par exemple.
RÉACTIONS PRINCIPALES ET MÉTHODES PRÉPARATRICES DE LA LAMELLE PORTE-OBJETS
AMYLOIDE
sur exsiccata
1) réhydrater le fragment à observer dans du KOH 5% pendant 1’ environ
2) effectuer un ou deux lavages dans l’eau et faire sécher
3) ajouter une goutte de Melzer et attendre 1’ environ
4) laver avec de l’Hydrate de Chloral et essuyer
5) Observer dans l’eau ou dans l’Hydrate de Chloral ou bien dans du L4 Tampon à la glycérine
sur de la matière fraîche :
1) exclure le passage dans le KOH 5%
2) procéder comme pour le point 3
CYANOPHILIE
sur exsiccata
1) réhydrater dans du KOH 5% pendant 1’ environ
2) laver la préparation 1-2 fois dans de l’eau et faire sécher
3) ajouter une goutte de Bleu lactique
4) faire réchauffer la préparation à feu doux pendant quelques secondes ; ajouter encore du Bleu lactique si nécessaire
5) retirer le fragment et le transposer sur une autre lamelle porte-objets
6) ajouter de l’Hydrate de Chloral
7) observer
NOTE : pour certaines genres comme Galerina, Rhodocybe ou Lepista n’est pas nécessaire de réchauffer.
CARMINOPHILIE OU SIDÉROPHILIE
sur exsiccata
1) réhydrater le fragment dans du KOH 5% pendant 1’ environ
2) laver la préparation 1-2 fois dans de l’eau et faire sécher
3) ajouter du Carmin Acétique
4) faire réchauffer le tout à feu très doux en remuant avec un bout de fer (pas d’acier) pendant quelques secondes
5) transposer la préparation sur une autre lamelle porte-objets et ajouter du Carmin Acétique ou bien de l’Hydrate de Chloral
6) écraser la préparation très fortement et observer
Table de travail © Pierluigi Angeli
RÉACTIFS PRINCIPAUX ET PRÉPARATIONS CHIMIQUES POUR LES OBSERVATIONS MICROSCOPIQUES
AMMONIAQUE (NH3) : on l’utilise pour la réhydratation des échantillons séchés (observation des exsiccata) ; normalement dans une solution aqueuse concentrée à 2-3 ou 6%.
HYDRATE DE POTASSIUM (KOH) : possède les mêmes propriétés que l’ammoniaque, on l’utilise donc sous forme d’exsiccata à une concentration maximale de 5%.
HYDRATE DE SODIUM (NaOH) : à l’identique des précédents, il faut l’utiliser à une concentration qui n’est pas supérieure à 5-10%.
HYDRATE DE CHLORAL (C Cl3 CH(OH)2) : sa propriété est celle d’éclaircir.
Dans le cas où l’on voudrait observer longuement la préparation, il faudra utiliser des liquides qui suppriment la possibilité de reproduction des microbes à l’intérieur de la lamelle. Le LACTOPHÉNOL D’AMMAN (solution d’Amman) composé de Phénol, d’Acide Lactique et de Glycérine, dans une solution aqueuse à 20%, est un liquide bactéricide très approprié, surtout dans le cas de longues observations pour effectuer des photomicrographiques ; il est important de bien ajuster la lamelle couvre-objet à la lamelle porte-objet (lutage : obtenu en posant un petit poids sur la lamelle couvre-objet et en en scellant les bords avec du vernis à ongles), de façon à empêcher d’éventuels débordements de liquide ou les infiltrations d’air.
Les solutions ammoniacales et la potasse sont utilisées également pour l’observation des matières fraîches, en effet chez certains genres (Galerina, Inocybe, Cortinarius), celles-ci colorient les parois des cellules marginales (kystiques) en provoquant une réaction importante d’un point de vue taxonomique.
ROUGE CONGO : c’est le colorant le plus diffusé en Mycologie, il teint les parois des cellules, s’adapte bien à de nombreuses structures cellulaires ; il est excellent même pour travailler avec les exsiccata lorsqu’il est employé dans une solution ammoniacale.
FLOXINA : il possède les mêmes propriétés que le Rouge Congo, il colorie le cytoplasme, est utilisé le plus souvent par les mycologues américains, on le trouve aussi bien dans les solutions aqueuses qu’ammoniacales.
BLEU COTON ou BLEU DE MÉTHYL : c’est le Bleu Coton au Lactophénol qui est le plus adapté, on l’utilise aussi comme réactif pour identifier la cyanophilie sporale ce qui veut dire que la membrane sporale tourne à une couleur intense (genre Entoloma) ou qu’elle ne se colorie pas (genre Tricholoma).
RÉACTIF DE MELZER : est obtenu à partir d’une solution d’Iode à l’Hydrate de Chloral, il sert principalement à vérifier l’amyloïde des spores. La réaction peut être amyloïde (membrane sporale de couleur gris-noirâtre ou bleuâtre, sommet des asques vert-bleu), dextrinoïde ou pseudo-amyloïde (la spore devient rougeâtre), inamyloïde.
CARMIN ACÉTIQUE : il est utilisé pour l’observation d’une réaction particulière qui a lieu à l’intérieur des basides des espèces qui appartiennent au groupe du Lyophyllum s.l. Les granulations de ces basides sont définies carminophiles ou sidérophiles et, typiquement, leur couleur tourne au gris-noirâtre.
NÉO-FUCHSINE BASIQUE (Fuchsine de ZIEHL) : cause une réaction sur les incrustations hyphales de la cuticule de certaines Russula, en acquérant une valeur taxonomique importante.
BLEU DE CRÉSYL : cause une réaction dite métachromatique dans les hyphes de certaines espèces du genre Mycena . La réaction se manifeste en teignant la paroi hyphale extérieure d’une couleur rouge-violette vive.
SULPHOVANILLINE : met en évidence les hyphes laticifères et les cystides des genres Russula et Lactarius .
SULPHATE DE FER (FeSO4) : est utilisé comme réactif macrochimique pour le genre Russula .
TEINTURE DE GUAIACUM : c’est aussi un réactif macrochimique très utilisé pour le genre Russula .
MENSURATIONS: QUOI, COMMENT ET COMBIEN
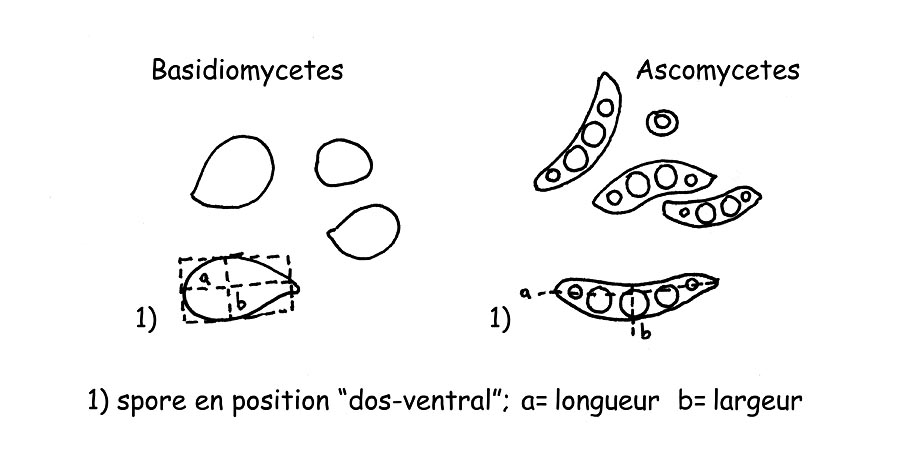
Mesure des spores © Maria Tullii
Spores: Pour effectuer la mensuration des spores on mesure normalement leur longueur et leur largeur en supposant qu’elles aient une forme géométrique hypothétique (un carré, un rectangle ou un cercle), en excluant leurs ornements et leur appendice hilaire. Les spores doivent être mesurées lorsqu’elles se trouvent en position ¨dorso-ventrale¨, comme dans le dessin (A). Normalement, on effectue environ 32 mensurations des spores des carpophores mûrs, préférablement sur la sporée. Pour une exposition correcte et utile des mesures prises il faudra calculer aussi bien les ¨Q¨ que les “QM” (soit le quotient et le quotient moyen) que le Volume ¨V¨ et le volume moyen “VM” des spores.
Pour trouver le “QM”:
il faut éliminer les mensurations qui sont manifestement anormales, aussi bien si elles sont trop petites que trop grandes, puis on trouvera le quotient de chaque mensuration, (ex. 10 x 6,5 – 10,5 x 7 – 9,5 x 7 etc. 10/6,5 = 1,53846153 qu’on arrondira à 1,53 + 1,5 + 1,35 etc. = 4,38 pour le diviser ensuite par le numéro de mesures avec lesquelles on a calculé le quotient on obtiendra le “QM”, pour cet exemple 4,38/3 = QM 1,46 ;
Pour trouver le ¨Q¨ :
on considérera parmi les quotients obtenus pour le “QM” le plus petit et le plus grand d’entre eux comme indication de mesure des spores Q= 1,35-1,53.
En ce qui concerne le volume des spores on procède de la manière suivante :
on relèvera la plus petite spore de notre champignon et on en calculera le volume à l’aide de la formule suivante V= 4π/3 × (a/2)² × b/2 (a= diamètre plus petit, b= diamètre plus grand).
on relèvera la plus petite spore et on en calculera le volume comme pour la précédente V1= 4π/3 × (A/2)² × B/2 (A= diamètre plus petit B= diamètre plus grand) on continuera ensuite avec la formule VM = (V + V1)/2, par exemple: dans une série de mensurations, la spore la plus petite mesure 6,5 × 4 µm et la plus grande 8 × 5,5 µm en appliquant la formula nous obtiendrions: V = (4×3,14/3) × (4/2)² × 6,5/2 = 54,34; V1 = (4×3,14/3) × (5,5/2)² × 8/2 = 126,40; VM = (54,34 + 126,40) / 2 = VM 90,4.
Basides et Cystides : On mesure la longueur à partir du point d’attache de la baside ou de la cystide jusqu’au subhyménium. On mesure la largeur au niveau du point médian ou au niveau de la largeur maximale.

Amanita muscaria – Amanita phalloides – Astraeus hygrometricus

Astraeus hygrometricus – Boletus aestivalis – Boletus edulis – Suillus granulatus

Tricholoma columbetta – Suillus granulatus – Tuber melanosporum – Tuber magnatum – Tuber melanosporum – Macrolepiota procera

Macrolepiota procera – Russula vesca – Otidea onotica

Verpa bohemica – Morchella esculenta – Marasmius oreades – Mutinus elegans – Macrolepiota procera

Leucoagaricus americanus – Amanita citrina – Amanita muscaria

Amanita muscaria – Amanita rubescens – Boletus aereus

Boletus aereus – Laccaria amethystina – Cortinarius rufoolivaceus

Cortinarius rufoolivaceus – Gyromitra infula – Cortinarius lividoviolaceus

Cortinarius lividoviolaceus – Amanita spissa


Amanita spissa – Cantharellus cibarius – Amanita mairei

Amanita mairei – Clitocybe gibba

Calvatia excipuliformis – Calvatia craniiformis – Clavariadelphus pistillaris

Clavariadelphus pistillaris – Clitocybe nebularis

Cortinarius turmalis – Clavulina cinerea – Gomphidius glutinosus

Gomphidius glutinosus – Clitocybe odora

Russula atropurpurea – Calvatia utriformis – Crucibulum laeve

Lactarius deliciosus – Suillus grevillei

Suillus grevillei – Russula mairei – Leccinum aurantiacum

Leccinum aurantiacum– Mycena inclinata – Oudemansiella mucida

Oudemansiella mucida – Mycena renatii – Lepiota clypeolaria – Lactarius torminosus

Lactarius torminosus – Badhamia utricularis

Badhamia utricularis

Clavulina rugosa

Russula foetens – Psathyrella caput-medusae – Cantharellus lutescens – Suillus luteus

Suillus luteus – Amanita muscaria – Stropharia aeruginosa – Tricholoma saponaceum


Amanita pantherina – Aleuria aurantia

Boletus aestivalis – Entoloma lividoalbum

Boletus edulis – Boletus aestivalis

Chroogomphus rutilus – Calocera viscosa – Gloeophyllum sepiarium

Stereum hirsutum – Lactarius aurantiacus – Fuligo septica

Fuligo septica

Fuligo septica – Cortinarius variecolor – Fistulina hepatica

Fistulina hepatica – Galerina marginata – Hypholoma sublateritium

Hypholoma sublateritium – Hygrophoropsis aurantiaca – Hypholoma fasciculare

Hypholoma fasciculare – Ischnoderma benzoinum

Ischnoderma benzoinum – Leucocortinarius bulbiger – Leccinum holopus – Spathularia flavida

Spathularia flavida – Paxillus involutus

Clitocybe costata – Fomitopsis pinicola – Lycoperdon perlatum

Lycoperdon perlatum – Lepista flaccida
= Lepista inversa – Suillus grevillei – Russula integra

Russula integra – Lactarius intermedius – Oudemansiella radicata

Oudemansiella radicata – Suillus granulatus – Clathrus ruber

Clathrus ruber – Tricholoma acerbum – Russula xerampelina

Ramaria lacteobrunnescens – Armillaria mellea = Armillariella mellea


Amanita vaginata – Amanita phalloides

Clitocybe costata – Boletus aestivalis – Boletus regius


Boletus edulis – Agaricus silvicola – Clitocybe candicans – Amanita rubescens

Cantharellus cibarius – Chroogomphus rutilus

Calocera viscosa – Clavulina rugosa

Cortinarius anomalus – Cortinarius cliducus – Cortinarius infractus

Cortinarius olivascentium – Cystolepiota seminuda

Cribraria argillacea – Ceratiomyxa fruticulosa

Ceratiomyxa fruticulosa = Famintzia fruticulosa

Ceratiomyxa fruticulosa var. porioides

Clavulina coralloides

Lactarius piperatus – Coprinus micaceus

Collybia fusipes – Cortinarius cotoneus – Dacrymyces abietinum

Ife miceliari

Ife miceliari – Hemimycena gracilis – Hemimycena pseudolactea


Hypholoma fasciculare – Hebeloma edurum

Lycoperdon perlatum – Lactarius deliciosus

Lactarius salmonicolor – Lactarius volemus

Lactarius porninsis – Lactarius serifluus – Leccinum aurantiacum

Lycogala epidendron

Lycogala epidendron

Lactarius circellatus – Lactarius intermedius – Laetiporus sulphureus – Hypholoma sublateritium – Leccinum carpini

Leucopaxillus candidus – Macrolepiota mastoidea

Lycoperdon echinatum – Lycoperdon molle

Macrolepiota procera – Mycena pura

Phaeomarasmius erinaceus – Merulius tremellosus

Phellodon tomentosus – Pluteus cervinus

Polyporus brumalis – Paxillus involutus

Russula cyanoxantha – Russula risigallina

Russula virescens – Russula aurea

Russula integra – Russula velutipes = Russula aurora

Russula delica – Marasmius oreades – Marasmius bulliardii – Scleroderma citrinum

Sowerbyella imperialis

Stropharia caerulea – Suillus granulatus – Trametes versicolor – Penicillum digitatum

Stemonitis axifera : phase juvénile

Stemonitis axifera : juvénile, en croissance et adulte
Stemonitis axifera adulta

Xerocomus subtomentosus – Xerocomus chrysenteron – Leccinum scabrum

Amanita rubescens – Amanita lividopallescens
Boletus satanas

Boletus satanas – Pholiota squarrosa
Ramaria formosa
Ramaria formosa – Tricholoma portentosum

Boletus edulis en âge mûr – Crucibulum laeve

Lasiochlaena benzoina = Ischnoderma benzoinum – Lycoperdon echinatum

Trametes versicolor – Trametes gibbosa

Phaeolus schweitnizii – Coprinus niveus

Cortinarius herculeus – Kretzschmaria deusta = Ustulina deusta : la forme grise est l’Anamorphe, celle noire le Téléomorphe

Inocybe geophylla – Tricholoma terreum – Perenniporia narymica – Laccaria bicolor

Stropharia caerulea – Coprinus comatus

Coprinus comatus – Amanita caesarea – Spore di Tuber melanosporum

Spores de Boletus edulis – Otidea umbrina – Tuber magnatum
© Giuseppe Mazza