

de Pierluigi Angeli y Maria Tullii

Traducción en español de Susana Franke
BREVE TRABAJO SOBRE LA HISTORIA DE LOS HONGOS

Agaricus praeclaresquamosus © Giuseppe Mazza
La micología es la ciencia que estudia los hongos. MICOLOGÍA, del griego MÝCHÊS = hongo y LÒGOS = discurso, tratado: micología quiere decir literalmente tratar sobre los hongos, micólogo es aquel que estudia los hongos.
Para hablar de hongos y entender ciertos comportamientos y modos es útil, a veces, partir desde tiempos muy lejanos: de la prehistoria.
Basándonos en testimonios hallados (grafitos) se puede suponer que el hombre haya observado los hongos desde tiempos remotos, y haya aprendido a utilizarlos, primeramente como alimento, experimentando la peligrosidad sobre su propia piel para luego de haber confrontado algunas propiedades particulares de ciertas especies, como hongos alucinógenos, usado en particular por brujos y chamanes, o como curativos, por la capacidad antiinflamatoria y para reforzar el sistema inmunitario. En tiempos relativamente recientes los hongos mortales ha sido a menudo utilizados para eliminar enemigos y personajes molestos.
Come se puede fácilmente imaginar al inicio los envenenamientos a través de hongos eran muy frecuentes y la dificultad de entender la causa hizo que nacieran supersticiones y falsa creencias, que aún hoy sobreviven en muchos lugares. Se pensaba que los hongos, inocuos por su naturaleza, se tornaran tóxicos a causa de factores externos, o bien se veía en ellos una creación del diablo para atraer al hombre y luego perjudicarlos.

Amanita pantherina © Giuseppe Mazza
Esta concepción del hongo puede ser trasladada a la manera de nacer y crecimiento del hongo mismo: éste surge de la tierra aparentemente de la nada (la planta hongo se desarrolla bajo tierra, no produce “semillas”, en la acepción común del término, y el crecimiento de los carpóforos no está estrechamente ligada a un ciclo estacional exacto, siguiendo las condiciones climáticas, variables de año en año), además el crecimiento de los carpóforos se desarrolla a menudo en círculos o líneas en zigzag, que venían cambiados por designio del maligno.
La prueba que algunas especies eran usadas para provocar alucinaciones y estados de trance en ritos chamánicos o adivinatorios, lo encontramos en muchos graffitis y diseños encontrados en varias partes del mundo: en el desierto del Sahara, en el territorio de Argelia, en el Valle de las Maravillas en Francia, en el estado indio, en el norte de Europa; por todas partes las figuras representaban ritos en grupo y los hongos representados hacen pensar en el género Psylocybe o en hongos seguramente con poder alucinógeno.
El uso adivinatorio no era el único que se le daba a los hongos: tenemos un testimonio reciente: en 1991 sobre el glaciar del Similaun, que divide Italia con Austria, se ha encontrado el cuerpo de un hombre muerto hace más de 5000 años y momificado naturalmente. Enrte los objetos que hacían parte de sus pertenencias, se encontró también Fomes fomentarius y Lenzites betulinus , el primero servía para encender el fuego, el segundo era usado en decocción para aumentar la resistencia física a la fatiga y al dolor, en polvo como antibiótico y hemostático para curar las heridas y también, probablemente, como cepillo para limpiar la piel.

Il Fomes fomentarius servía en la antiguedad para encender el fuego © Pierluigi Angeli
Aún hoy, en los países escandinavos, el Lenzites betulinus, considerado un vigorizante natural, viene consumido en decocción. En tiempos más recientes, se comienzan a encontrar, si bien no en cantidad, testimonios escritos sobre hongos, su naturaleza, su utilización, las creencias ligadas a ellos. Uno de los primeros escritos documentados sobre los hongos en una obra de la antigua Grecia habla como se cultivan “… enterrar excrementos equinos debajo de un árbol de higo y regarlo abundantemente…”. El griego TEOFRASTO (370 – 287 a.C.) nativo de la isla de Lesbos y discípulo de ARISTÓTELES, que es considerado el padre de la botánica, habla así de los hongos: “… ellos son plantas imperfectas, sin raíces, sin hojas, sin flores y sin frutos”.
Después de TEOFRASTO el tema de los hongos pasó a la esfera de competencia de los médicos y es necesario llegar al 50 d.C. para encontrar otra vez escritos sobre hongos.
En la obra “DE MATERIA MEDICA”, PEDACIO DIOSCÓRIDES, nacido en Cilicia (Asia Menor), médico militar y civil en Roma bajo los emperadores Claudio y Nerón, nos trasmite algunas nociones sobre la toxicidad de los hongos y sobre la terapia a seguir en caso de envenenamiento: “… decocción de hierbas aromáticas, pociones de vinagre y sal, suministrar estiércol de pollo formando una pasta con miel y vinagre …”, remedios que probablemente han sido usados por muchos siglos. A él se debe la primera descripción del AGARICUM (Laricifomes officinalis) que era usado como medicinal.

Boletus satanas © Giuseppe Mazza
Otro médico que se ocupó de hongos, griego de Pérgamo; que vivió en Roma en los tiempos de los emperadores Marco Aurelio, Lucio Vero y Cómodo; fue CLAUDIO GALENO (129-200 d.C.). Galeno había dividido a los hongos en tres grupos fundamentales: “Bolites”, “Amanitai” y “Mikés”. Al primer grupo pertenecían los hongos con laminillas (entre los cuales las actuales Amanitas, género al cual pertenece Amanita caesarea que aún hoy, en algunos lugares de Italia, es llamada comúnmente “boleto”); al segundo grupo todos los hongos con tubos y esporas (el actual género Boletus, según la nomenclatura creada por Linneo en 1735); al tercer grupo finalmente correspondían todos los otros hongos con sombrero y pie. Según Galeno los hongos pertenecientes a los primeros dos grupos se consideraban todos comestibles, mientras aquellos del tercer grupo se consideraban tóxicos y no aptos para el consumo, porque “… no nutritivos, difíciles de digerir y peligrosos por las propiedades tóxicas …”. Galeno finalmente confirmaba la validez de la terapia de estiércol de pollo contra los envenenamientos.
El primer autor que trató los hongos con más profundidad, ha sido Cayo Plinio Segundo llamado PLINIO EL VIEJO (23 – 79 d.C.). Él nos ha dejado una obra en 37 libros “HISTORIA NATURALIS” donde, entre otros argumentos naturalísticos, habla también de hongos. He aquí algunos extractos: “… entre las comidas menos recomendables se deben mencionar los boletus …” (los actuales Amanitas ndr) y motiva su afirmación con la facilidad de confundirlos con hongos tóxicos.

Amanita muscaria © Giuseppe Mazza
Refiriéndose a Amanita muscaria dice “… algunos se distinguen por una suerte de verrugas que están sobre el sombrero, blancas y secas, similares a florescencias de sal nitro, y que no son más que residuos del velo …”.
Además cita algunas circunstancias en donde hongos perfectamente comestibles pueden transformarse tóxicos y dice: “… Si nacen cerca de botones metálicos, clavos de zapatos, hierros herrumbrados, paños húmedos, absorbiendo los jugos impregnados de tales sustancias y transformándolas en veneno”.
Y algo más: “… si en las cercanías hay alguna cueva de serpiente, o si una serpiente en su camino les sopla encima, el hongo se transforma en venenoso, porque su naturaleza es de absorber cualquier tipo de sustancia venenosa”.
Estas creencias están aún fuertemente presentes y tenidas como veraces en casi todo el territorio italiano. Se creía también que la comestibilidad de los hongos se podía intuir observando las plantas que los rodeaban. Plinio definía inocuos los que nacen debajo de las coníferas, el higo y la cañaheja; tóxicos aquellos que nacen debajo de los robles, hayas y cipreses.

Amanita phalloides © Giuseppe Mazza
Esto se debe probablemente a que las Amanita mortales, y la phalloides en particular, prefieren las latifolias, mientras que sabemos que sobre las raíces de la cañaheja nace el Pleurotus eryngii, óptimo hongo conocido desde la antigüedad.
Plinio describe también el desarrollo de los boletus: “… nacen de la tierra, primeramente todos envueltos en su volva, que lo hace asemejarse a un huevo. En tal estadio de desarrollo estos hongos son particularmente apreciados. A continuación la volva, rompiéndose, queda adherida a la base del pie …”.
Por primera vez es usado el término volva en su correcto significado micológico. Él describe luego la duración de un boletus que es de sólo siete días, el crecimiento no cespitoso, con carpóforos no en pares sino simples.
Para explicar la naturaleza de los hongos (que nacen aparentemente de la nada) Plinio dice: «… Su origen hay que buscarlo en el fango de la tierra húmeda y de sus estados, que empiezan a fermentar, o en las raíces de las plantas cupulíferas ….»
Esta afirmación quedó radicada por siglos en las opiniones de los naturalistas, hasta que, con las nuevas instrumentaciones, se logra dar una connotación más científica en el estudio de la materia. Del boleto Plinio hace un hongo noble y lo trata aparte, mientras relega a todos los otros juntos y dice: «… otros hongos tienen una consistencia más dura. De ellos existen infinitos géneros, pero todos tienen un origen en común: la linfa de los árboles. Son comestibles segurísimo aquellos de color rojo más oscuros que los boletus …».
En este escrito habla, con mucha probabilidad, de hongos lignícolas y sin ninguna duda de Fistulina epatica que era considerada óptimo comestible desde tiempos antiquísimos.

Fistulina hepatica © Giuseppe Mazza
Dice además: ”… hay también especies de color blanco, con pie muy desarrollado y sombrero que se parece al sombrero de un Flamen …”. El Flamen era un sacerdote de una antigua religión romana que tenía un sombrero que en su aspecto recordaba a un Macrolepiota procera con sombrero no del todo abierto.
Dice además: “… un tercer género de hongos, los Suilli está muy relacionado con el veneno…”.
Los “Suilli” eran los actuales Boletaceae, y eran considerados venenosos por el color que cambia a lívido, o sea azulino. Al contrario, en tiempos de Plinio los Suilli eran un producto muy comercializado: se importaban de Asia Menor y colocados ensartados en un junco delgado y puestos a secar cerca del fuego, sistema usado aún hoy en algunos lugares de nuestro país (Italia). Además algunos Suilli eran usados por los médicos para curar hemorroides, granos, mordeduras y también para prepara un colirio.
Pero el peligro mayor, afirmaba Plinio, venía de los porcini tóxicos que han hecho estragos en enteras familias. Plinio, en su tratado, aconseja: “… si alguien quiere realmente comerlos, trate de prestar atención en el momento de la compra y controle particularmente su cocción …”. Y concluye: “… desconfíen de aquellos hongos que al cocinarlos endurecen, se hagan cocinar mucho tiempo, poniendo en el recipiente también salitre. Se cocinen preferiblemente junto a la carne (para diluirlos) y con pecíolos de peras selváticas (diversamente éstos últimos podrán ser comidos al finalizar la comida como antídoto).

Macrolepiota procera © Giuseppe Mazza
Se tenga presente que el aceto es por su naturaleza contrario a los venenos, y por lo tanto se usa siempre sin moderación cuando se comen los hongos ”. De resaltar como este último consejo si se aplica en la preparación de Coprinus atramentarius, puede provocar una intoxicación seria como el síndrome coprínico.
DEL MEDIOEVO AL SETESCIENTOS
Con la caída del imperio romano de occidente y la consiguiente barbarie de las poblaciones, debido a las invasiones de los pueblos nórdicos, se entra en aquello que es llamado “medioevo”. La cultura clásica y científica pasa a las manos del clero y sobre todo de los monjes Benedictinos que conservan las obras, las transcriben y las traducen. El medioevo es mezquino en noticias y pocos son los estudiosos de hongos que nos han transmitido sus obras. Es sin embargo con el resurgimiento que se abre una nueva era para la botánica, que es reconocida como verdadera ciencia, aunque sólo como complemento de la medicina. En este período vienen desempolvados Dioscoride y Plinio con traducciones, comentarios y reediciones
ERMOLANO BARBARO (1454-1492) amante de la cultura de Aristóteles, Plinio y Dioscoride, tradujo y comentó sus obras.
PIER ANDREA MATTIOLI (1500-1577)
El Mattioli, nacido en Siena pero trentino por adopción, ha sido un médico de gran fama. Su obra principal es Commentari in Pedacii Dioscoridis Anarzabei de Materia medica,en ella viene reportada la traducción del griego al latín de la obra original de Dioscoride, En cada capítulo ha inserido como apéndice, un largo comentario mucho más extenso que el capítulo mismo, Fue una obra de gran valor tal es así que en dos siglos tuvo 60 ediciones, en original, en italiano, en francés, en alemán y en checo. En aquella época, cuando la comunidad científica comenzó a considerar a la botánica como una verdadera disciplina, aunque ligada a la medicina, los hongos venían tratados marginalmente.

Cordones miceliarios en el subsuelo © Giuseppe Mazza
Sin embargo el 1500 ha visto el nacimiento de los primeros tratados monográficos. El primero de ellos ha sido publicado por un médico holandés: Jonghe Hadrian (1512-1575) que en 1564 escribe una disertación sobre una nueva especie que llamó Phallus impudicus , con el título “Phalli ex fungorum genere in Hollandiae sabuletis passim crescentis, descripto et ad vivum expressa pictura”, donde estaba descripto con una tabla xilográfica, un hongo, que seguidamente vendría llamado en su honor Phallus hadriani ; que crecía en los terrenos arenosos en Holanda. El opúsculo reportaba también las virtudes medicinales del hongo, considerado terapéutico para la gota y dolores artríticos
PIER ANDREA CESALPINO (1525-1603) nacido en Arezzo en 1525, casi contemporáneo con el Mattioli, también el médico celebérrimo, el Cisalpino ha dejado una profunda impronta de si mismo en la botánica. En campo médico fue el primero en descubrir y describir en términos modernos la circulación sanguínea y los capilares sanguíneos, aunque si el mérito fue atribuido, injustamente, mas tarde al inglés Harvey.
El Cisalpino ha sido médico del Papa Clemente VIII, fue arquetipo pontificio y profesor de medicina en la Sapienza de Roma, ha sido médico y amigo de San Filippo Neri y Torcuato Tasso. Su principal trabajo de botánica ha sido la obra De Plantis en 16 libros, donde pone en acto un primer intento de catalogar las plantas de manera sistemática, según criterios precisos. Por esto es considerado el padre de la botánica italiana. El Cisalpino, como botánico, trata obviamente también los hongos, es el primero que intenta hacer un estudio profundizado sobre ellos, y es el primero que mas allá de las descripciones de las varias especies, da consejos sobre como cocinar los comestibles. Hasta aquí, el conocimiento de los hongos, se puede decir, no había hecho grandes progresos. Pesaba fuertemente la influencia de las antiguas creencias, la ausencia de instrumentos científicos aptos dejaba vacíos infranqueables sobre todo sobre las modalidades de su nacimiento, se creía que los hongos fueran formas patológicas de las raíces o de los árboles, cuando no incluso que tuvieran orígenes mágicos. El progreso de la botánica había sido notable, mucho menos el de los hongos.
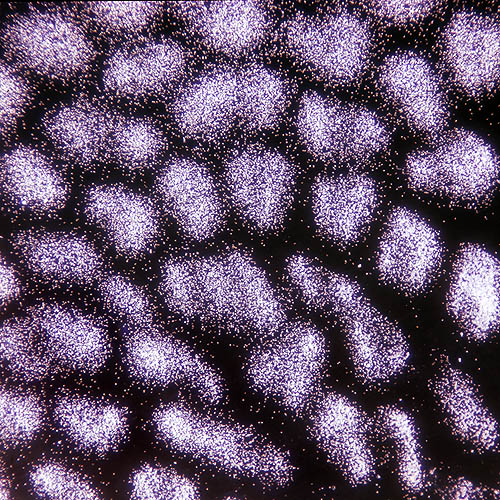
Esporas di Boletus aestivalis en el microscopio, con débil ampliación © Giuseppe Mazza
DESDE EL SETESCIENTOS HASTA NUESTROS DÍAS
Con el setecientos han sido diversos científicos que han hablado de hongos y han intentado darle a ellos una sistemación en el mundo de la botánica, pero es con la llegada de PIETRO ANTONIO MICHELI (Firenze 1679-1737) que nace la Micología moderna, así como aún hoy la conocemos.
En Micheli la pasión por la botánica nace de la literatura del Mattioli, del Bocón y de otros científicos, mientras trabajaba como joven empleado de tienda junto a un encuadernador de libros. Se convierte en un botánico de fama internacional, aún sin tener ningún título de estudio, empujado sólo por una gran pasión, ayudado por un erudito y también por los monjes benedictinos de Vallombrosa (Firenze), en particular, en cuanto se refiere a la micología, sobre todo por el Padre Bruno Tosí, gran botánico de la época, socio de la Royal Society de Londres.
El Micheli, es considerado, con justo mérito, el padre de la micología moderna. A el se le atribuye el gran mérito de haber descubierto y demostrado que también los hongos se reproducen por semilla, con el descubrimiento de las esporas y su germinación. Firme era su convicción de que ninguna planta se pudiera reproducir sin semilla, y que no existe ninguna planta privada de flor y semilla. Con el auxilio de un rudimentario microscopio de tres lentes, donde para aumentar las ampliaciones era necesario cambiar físicamente el objetivo, el Micheli logró descubrir y describir los cheiloscytidia y los pleurocystidia. Así describe lo que ve:
“ … diminutos semolines distribuidos debajo de ellos, por cuanto observar pude, con orden regularísimo; y aquello q ue me ha hecho maravillar aún mas ha sido el ver que cada uno de ellos estaba situado sobre una base, lo cual me hace dudando decir: quien sabe que no sean la flor o el cáliz de los hongos ? ”
Algo más:
“… en el borde de la lámina… nacen flores apétalas, desnudas, constituidas por un simple filamento cilíndrico; en algunas especies estas flores son solitarias, o si no unidas, mientras en otras especies están reunidas en masa o copos. Sobre la superficie de la lámina nacen por todas partes semillas redondas o esféricas, diminutas, en algunas especies dispersas singularmente, en otras especies reunidas cuatro por cuatro”.
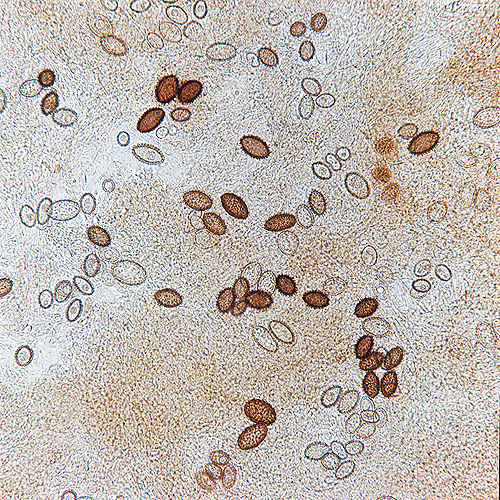
Esporas de Tuber melanosporum observadas en el microscopio © Giuseppe Mazza
“ …en algunas especies de hongos, y particularmente en aquellos que nacen sobre el estiércol equino, bovino y similares, hemos visto algo digno de consideración: y es que sobre la cara de las láminas se encuentran no solamente las semillas sino también ciertos cuerpos diáfanos, de forma a veces cónica, a veces piramidal, y que parecen haber sido colocados allí providencialmente por la naturaleza para impedir que las láminas se toquen con aquello adyacente y obstaculicen, reteniéndolas entre ellas, la caída de semillas que alcancen la maduración …”.
La historia de esta fascinante materia se enriquece notablemente en el ‘800 y 900, también gracias a nuevas metodologías y nuevos instrumentos, pero nosotros nos detenemos aquí, porque sería largo contar toda la historia de la micología luego del Micheli. Es más, desde el setecientos en adelante hemos tenido grandes botánicos, sistemáticos y micológos, desde Linneo a Fries a Persoon y tantos más aún, hasta llegar a Don Giacomo Bresadola, el más grande micólogo desde fines del 800 e inicios del 900 quien, sin jamás moverse de Trento, ha estudiado los hongos de todo el mundo mediante las relaciones conseguidas con los más grandes científicos del globo.
Dedicaremos algunas líneas más para el gran Carl von Linné, nacido el 23 de mayo de 1707 en Rashult, en la provincia de Smaland, en Suecia, y muerto en Uppsala en enero de 1778, más conocido como Linneo.
En su obra más importante: “Systema naturae” (1735) illustra por primera vez su método de clasificación taxonómica de los reinos animales, vegetales y minerales. En 1753 con la obra “Species Plantarum” introduce la nomenclatura binomial donde cada animal, cada planta y cada mineral son indicados con dos nombres latinos, el primero indica el género mientras el segundo indica la especie.
La revolución derivada del sistema de Linneo hizo que en 1867, en París, se haya organizado el primer Congreso Internacional de Botánica donde fueron adoptadas las “Leyes de nomenclatura botánica”. Fue durante el segundo Congreso Internacional de Botánica realizado en Viena en 1905 que se adoptan las “Reglas internacionales de Nomenclatura Botánica” (el primer código) Desde entonces cada aproximadamente cinco años reanuncian Congresos a nivel mundial para aportar eventuales modificaciones. El último se ha realizado en Viena en el 2005 y en el 2006 ha sido publicado el último código Internacional de Nomenclatura Botánica denominado (Viena Code).
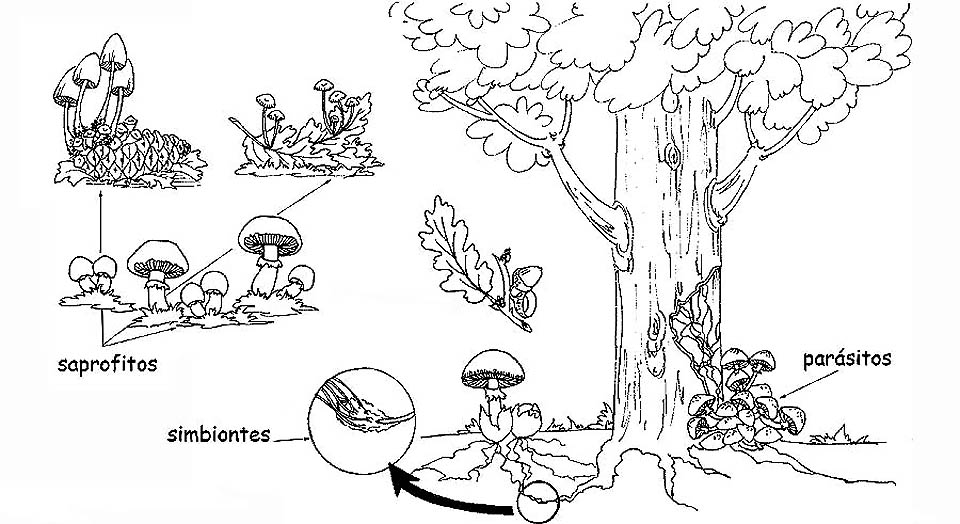
Modos de nutrición de los hongos © Maria Tullii
EL HONGO
El hongo es un organismo viviente formato por células unidas y comunicantes entre ellas, en una estructura constante, diversificadas entre ellos en función de su fin biológico. El verdadero organismo hongo está constituido por un conjunto de filamentos (llamados hifas) que se esparcen en el interior del sustrato, a menudo por varios metros. El conjunto de estas hifas se llama “micelio”. Singularmente las hifas no son visibles a simple vista pero se entrelazan entre ellas formando aglomerados miceliares: “cordones miceliares” o “esclerocio”. Las hifas se alargan en el terreno mediante la subdivisión de la célula apical o de células intermedias, que dan origen a ramificaciones laterales. De este modo el micelio aumenta la propia superficie y la propia capacidad de absorción de sustancias nutritivas. El hongo comúnmente dicho, aquel que es recolectado, no es otra cosa que el fruto producido por el micelio cuando las condiciones de humedad y temperatura son favorables. El fruto es más correctamente llamado “carpóforo” o “sporóforo”. También el está constituido por aglomeración de hifas. Como para todos los otros frutos su función es la de la reproducción, que veremos en detalle más adelante.

Ciclo reproductivo de las setas © Maria Tullii
EL HÁBITAT
Los hongos nacen y crecen prácticamente en cualquier hábitat. Pueden preferir las zonas abiertas, o los ambientas más húmedos, un terreno ácido o básico, la convivencia con esencia arbóreas o arbustivas o de praderas, resumiendo, cualquiera sea la conformación del hábitat habrá una o más especies de hongos aptas para colonizarlo. Además existen especies de hongos que llamaremos “ubicuitarios”, que están en grado de colonizar hábitat diversos. El hábitat, mientras tanto, influye sobre el crecimiento y sobre el aspecto de los carpóforos: la misma especie puede presentar dimensiones y caracteres morfológicos diversos según la altitud, de la mayor o menor humedad, de la mayor o menor exposición a los rayos solares.
CONDICIONES DE CRECIMIENTO
Las condiciones de crecimiento del organismo hongo están fuertemente dependiente de las condiciones ambientales: cuando humedad y temperatura son favorables el micelio produce los carpóforos, que se desarrollan y producen las esporas que, a su vez, cayendo en el terreno, darán inicio a un nuevo ciclo vital. Las condiciones de crecimiento varían de zona en zona: el período de crecimiento de los hongos será más breve allí donde el clima es más rígido y la estación invernal es prolongada, casi nulo en las regiones de carácter permanente de sequía, prácticamente ininterrumpido en las regiones de clima templado.
LA REPRODUCCIÓN
Como se decía, el micelio, en condiciones climáticas de humedad y temperaturas oportunas, fructifica, dando origen a los carpóforos. Para reproducirse, mediante células especializadas, producen millones de semillas llamadas “esporas” que, dispersadas en el ambiente circundante con el auxilio de factores externos como el viento, lluvia o vectores animales, también a gran distancia, dan origen a un nuevo ciclo vital. Las esporas son células pequeñísimas, no visibles al ojo humano, observables por lo tanto solo con el microscopio. La unidad de medida de las esporas es el micrón (µm), que corresponde a un milésimo de milímetro. Las esporas son sexuadas (al menos en los hongos superiores, que son a quienes estamos aquí tratando) no en el sentido común del término (masculino o femenino) sino por la diversa carga que poseen: positiva (+) o negativa (-).
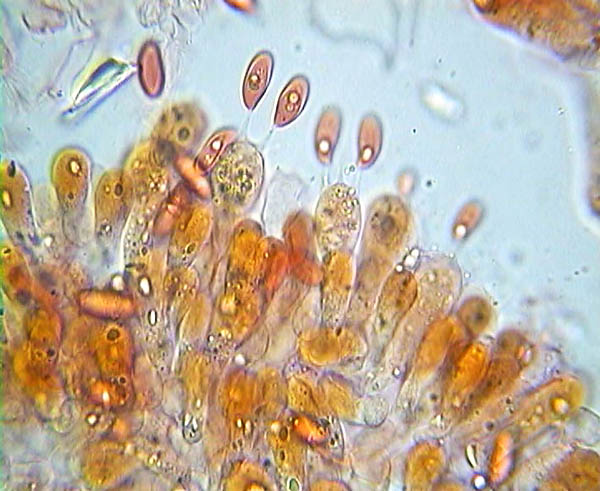
Basides et spores de Lepiota alba © Pierluigi Angeli
Cuando una espora encuentra un sustrato apto para la reproducción y en condiciones favorables, germina, dando origen a un filamento con la misma carga que la de la espora originaria (hifa), que puede desarrollarse y crecer (micelio primario), pero no puede fructificar.
Para que el micelio primario pueda fructificar, debe combinarse con otro micelio primario de la misma especie pero de signo opuesto.
La unión de los dos micelios, negativo y positivo, es llamado micelio secundario, el organismo “hongo” real y verdadero. No obstante, la posibilidad que del micelio primario se forme una nueva colonia de hongos es prácticamente rara y es por esto que cada cuerpo fructífero deja en el ambiente millones y millones de esporas.
SISTEMAS DE NUTRICIÓN
Todas las formas de vida presentes sobre la Tierra se dividen en:
PRODUCTORES o AUTÓTROFOS: organis- mos vivientes en grado de nutrirse de manera autónoma sintetizando las sustancias que tienen necesidad partiendo de los compuestos inorgánicos presentes en la naturaleza. Las plantas usan la luz solar para producir los compuestos complejos (como los azúcares) a partir de compuestos simples como anhídrido carbónico y agua, a través de el proceso de la Fotosíntesis clorofiliana. CONSUMIDORES o HETERÓTROFOS: organismos vivientes que se procuran las sustancias necesarias para su supervivencia de productores o de otros consumidores. Las fuentes de donde obtienen la comida, permiten subdividir a los consumidores en cuatro grupos: Herbívoros: se nutren de vegetales; Carnívoros: se nutren de otros consumidores; Omnívoros: se nutren tanto de sustancias vegetales como de otros consumidores; Descomponedores: se nutren utilizando los organismos muertos o vivientes, transformando las sustancias orgánicas complejas en sustancias simples, como anhídrido carbónico y agua. Las plantas superiores, siendo capaces de alimentarse a si mismas, transformando las sustancias inorgánicas (agua y anhídrido carbónico) en compuestos complejos, mediante el proceso de la fotosíntesis clorofiliana, pertenecen al grupo de los organismos Autótrofos. Los hongos, un tiempo clasificado entre los vegetales, pertenecen ahora a un reino propio: el reino Hongos, en cuanto no están en grado de desarrollar la función clorofiliana. Están por lo tanto para ser contados entre los organismo Heterótrofos y entre ellos en los Descomponedores.
En base al sistema nutricional adoptado, los hongos se clasifican en:

Clitocybe odora, hongo descomponedor de la hojarasca © Giuseppe Mazza
Saprófitos: se nutren de sustancias orgánicas en descomposición, ya sean vegetales o animales. Realizan una primera fase de disgregación y preparan el terreno a las bacterias que completarán el ciclo, de modo que todos los residuos orgánicos se retransformen en sustancias inorgánicas: agua, anhídrido y sales minerales. Hacen parte de este grupo los hongos que nacen sobre prados y pastos, hojas y ramas secas u organismos animales muertos. Podemos considerar a los hongos saprófitos como operador ecológico de la naturaleza: sin ellos los detritos vegetales y animales sofocarían el ambiente.
Parásitos: viven a expensas de otros seres vivientes, ya sean vegetales o animales. Si por una parte, los hongos parásitos representan un verdadero peligro para plantas y animales, porque provocan serias patologías hasta llegar a causarles la muerte, por otra parte, cumplen el rol de elemento regular, suprimiendo las plantas más débiles o enfermas, favoreciendo el desarrollo de aquellas sanas. Una vez muerto el organismo huésped, vegetal o animal que fuera, el hongo parásito se transforma en saprófito, terminando de demoler la sustancia orgánica muerta.
Micorrizas o simbióticos: son los hongos que viven en simbiosis con las plantas superiores, estableciendo una relación de mutua ventaja. Las partes apicales del micelio envuelven la parte terminal de los pelos radicales de las plantas superiores, aumentando la superficie de absorción. La planta tiene así a disposición una mayor cantidad de sustancias inorgánicas, que transformará, mediante la fotosíntesis clorofiliana, en compuestos orgánicos, que pone a disposición del hongo. Además la micorriza (formado por los pelos radicales envueltos de las hifa del hongo) protege las raíces de la planta de los ataques patógenos, haciéndola mas fuerte y mejorando la salud. Las micorrizas pueden ser: Endomicorrizas, cuando las hifas de los hongos penetran en el espacio intercelular y en el interior de las células vegetales. El hongo se expande a través de las células corticales, sin resquebrajar el cilindro central y las células apicales de la raíz. Ectomicorrizas, cuando las hifas de los hongos circundan las raíces penetrando en los espacios intercelulares pero no en el interior de las células vegetales. El manto de las hifas alrededor de las células radicales es llamado retículo de Hartig.
Homogeneidad del hongo © Maria Tullii
MORFOLOGÍA DE LOS HONGOS
Para que a un hongo se le pueda asignar un apellido y un nombre correctamente (Género, especie) es necesario observar todos los caracteres.
Comencemos a distinguir algunas tipologías de los hongos:
Agaricales son todos aquellos hongos compuestos de sombrero y estipe;
Ascomiceti: se presentan a veces con seudo estipe y seudo sombrero, o bien en forma de copa, sésiles o no, o bien en forma de tubérculos y en este caso, hipogeos o semihipogeos
Gasteromiceti en forma ± redonda, con o sin seudo estipe, hipogeos y epigeos.
Examinemos ahora los caracteres morfológicos que se deben observar en los hongos compuestos desde sombreo a pie: las diferencias, tal vez, son mínimas y sólo una observación cuidadosa nos consiente a exacta determinación.

Forma del sombrero © Maria Tullii

Margen y perfil del sombrero © Maria Tullii
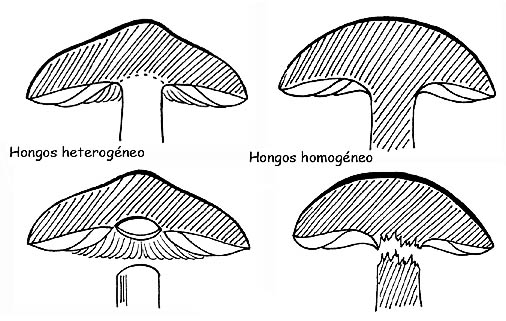
HOMOGENEIDAD
El carpóforo está constituido por un manto de hifas estrechamente unidas y dispuestas generalmente paralelas entre ellas. Si estas hifas tienen la misma estructura ya sea en el sombrero que en el estipe, tendremos un hongo homogéneo. Esta característica es fácilmente verificable: si cuando se intenta separar el sombrero del estipe, se rompe. Al contrario, si las hifas están estructuradas en modo diverso en el sombrero y en el estipe, tendremos un hongo heterogéneo y por lo tanto la separación de estas dos partes se podrá hacer sin roturas, como si las dos partes fueran simplemente apoyadas una sobre la otra.
SOMBRERO
Del sombrero se observan:
las dimensiones, correspondientes a su diámetro medido en su completo desarrollo.
la forma que cambia notablemente desde su estadio juvenil al maduro. Normalmente, se toman en consideración los ejemplares adultos, pero sería mejor observar más ejemplares, en diversos estadios de crecimiento. En lo ejemplares jóvenes, normalmente, el sombrero es más cerrado para luego abrirse y aplanarse hasta alcanzar en el adulto, la forma típica.
el margen, siempre importante, incluso determinante en algunas especies, si observa atentamente con referencia al plano horizontal del sombrero. Puede presentarse derecho y por lo tanto sobre el mismo plano; enrevesado, enrollado hacia adentro; revoluto enrollado hacia fuera; recurvo, plegado hacia abajo; El perfil del margen puede presentarse: regular, ondulado, lobado, festoneado, sinuoso o plisado, entero o estriado; es además considerado excedente cuando sobresale, aún poco, mas allá del himenio (láminas, tubos, etc.).

Decoraciones del sombrero © Maria Tullii
la cutícula del sombrero, puede tener diversos aspectos: lisa o viscosa, tomentosa, peluda, aterciopelada o áspera. Observamos, además, si se puede separa de la carne del sombrero: la cutícula que no se separa se llama adnata o no separable, si por el contrario, se separa enteramente, se dice que es separable, si se separa sólo en parte,
diremos que es parcialmente separable y observaremos la porción de rayo del sombrero que se descubre (1/2, 1/3, etc.).
la superficie del sombrero, puede o no presentar decoraciones. Una superficie in decoraciones unida, glabra, uniforme; donde están presentes las decora- ciones pueden ser:
· Innata, o sea parte integrante de la estructura de la película, como fibrillas, verrugas o machas.
· Adnatas o aplicadas, derivantes de las disociaciones de la película como craquelado y escamas, o bien residuos del velo general como verrugas, placas, harinosidad, vellosi- dad, pruinosidad.
el color es un carácter importante pero no absoluto, en cuanto la mayor parte de los hongos presenta una variabilidad cromática notable en relación a las condiciones climáticas, a la edad, a la exposición.
Para describir un color haremos referencia a colores comúnmente conocidos y asociados a objetos cotidianos (per ejemplo: rojo ladrillo, verde oliva, etc).
HIMENÓFORO
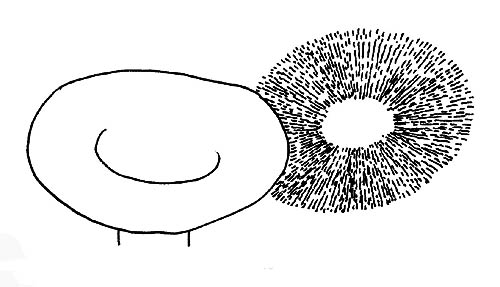
El himenóforo es el conjunto de las estructuras macroscópicas y microscópicas encargado a la producción de esporas.
Esporada en masa © Maria Tullii
Esto reviste importancia fundamental, no solo desde el punto de vista biológico, sino también de la determinación.
En los hongos con estipe y sombrero está generalmente situado en la zona inferior del sombrero y puede estar formado por láminas, tubos y esporas, punzadas; en algunos géneros es casi liso o apenas rugoso.
Expuesto al aire en la mayor parte de los casos, el himenio es en cambio protegido y encerrado en el interior del carpóforo en los hongos pertenec ientes a los Gastromycetidae (las esporas vendrán liberadas, en la maduración del carpóforo, por rotura del peridio) y, entre los Ascomycetes a los Tuberales .
Las láminas: el primer carácter que debemos observar es el color de las láminas, que puede ser innato, o sea el color visible es propio de la lámina; derivado, o sea el color de la lámina deriva de la coloración de las esporas.
Carácter este de enorme importancia en cuanto la pertenencia a un grupo cromosporeo, determina la asignación de un hongo a un grupo de géneros o a otro. Los grupos son cinco: leucospóreos, esporada blanca o blancuzca; ocrospóreos, esporada ocre; rhodospóreos, esporada rosa; ianthinospóreo, esporada violeta; melanospóreo, esporada negra.
Partes de la lámina © Maria Tullii
El color de la esporada es verificable a través de la recolección del polvo esporal sobre soporte de color contrastante.
Observaremos luego la homogenidad de las láminas, análogamente a todo lo ya dichopara el sombrero y por la cutícula observaremos si las láminas se separan fácilmente de la carne del sombrero: si es así diremos que son heterogé- neos o separables, si no se separan y se rompen en el intento, diremos que son homogéneos o no separables.
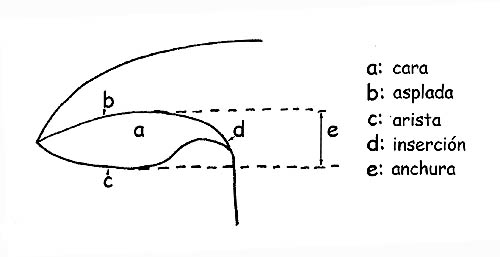
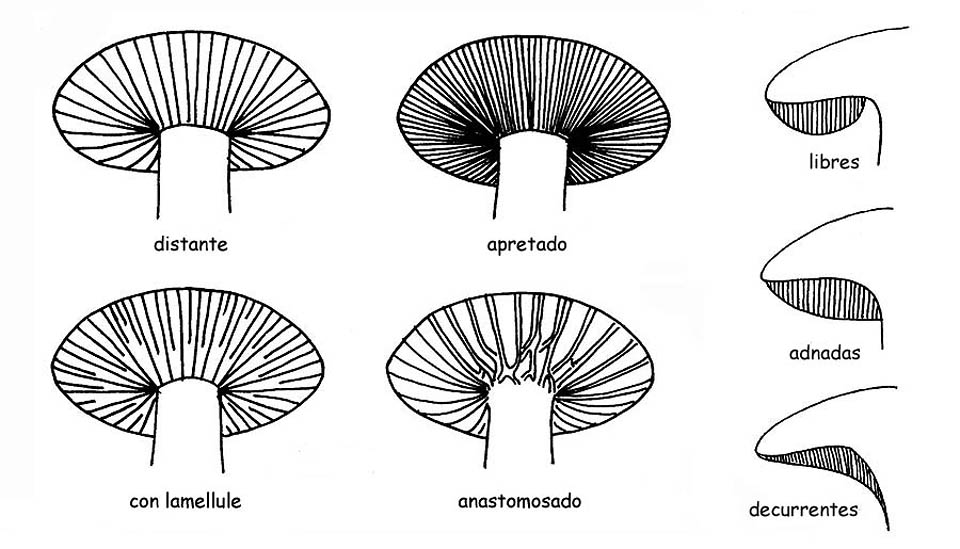
La forma de las láminas en todos sus componentes: orilla, hilo, largo, espesor, etc.; la inserción sobre el tronco, por lo que vienen definidas:
libres o distantes, cuando las láminas no legan a tocar el tronco, característica común a todo los hongos heterogéneos.
adnatas, cuando las láminas se pegan al tronco en todo su ancho.

Inserción de las laminillas © Maria Tullii
sin margen o uncinadas, cuando se unen al tronco sólo con una pequeña parte de su ancho.
decurrentes, cuando continúan a lo largo del tronco, mas allá de su ancho.
decurrentes por un dientecito, cuando la parte que decurre forma una pequeña prolongación (dientecito).
collarium, cuando las láminas, alejadas del pie, se unen a él mediante un pequeño cordón circular.
Observando el perfil laminar en el borde, podremos definirlas agudas, sinuosas, ventrudas, obtusas o derechas.
El dorso es el borde laminar unida a la carne del sombrero.

Perfil de las laminillas © Maria Tullii
El hilo laminar (que es el borde opuesto al dorso) puede ser entero, aserrado, dentado, liso, algodonoso, pruinoso y puede ser concoloro a las láminas o de color diverso.
En este caso hablaremos de hilo heteromorfo.
Si en cambio el hilo laminar se divide en proximidad del pie, las láminas se dicen furcados.
Presencia o menos de laméllas.
Están así definidas aquella láminas que parten del margen del sombrero y no llegan al pie.
Están normalmente intercaladas con las láminas y tienen largo variable.
La parte terminal de las laméllas puede ser trunca, redondeada o sinuosa, presencia o menos de Anastomosis.
Se trata de conjunciones transversales en más puntos una lámina con la otra.
Para mayor claridad por ancho se entiende la distancia entre el dorso y el hilo laminar.
Tipología laminillas y tubos © Maria Tullii
Con la definición espesa o sutil se refiere al espesor de la lámina.
Inserción del pie © Maria Tullii
Con las definiciones raras, densas se refiere a la distancia entre una lámina y la otra.
Análogamente a los caracteres indicados para las láminas, con los debidos ajustes, observaremos los caracteres del himenio en Tubos y poros.
Observaremos el color de los tubos y de los poros en el hongo joven y en el hongo maduro (tubos y poros pueden ser concoloros pero también de color diverso) con todas las variaciones intermedias; el largo de los tubos, la manera de cómo están unidos al tallo, el tamaño de los poros, su forma, etc.
Nota: En los himenóforos con tubos y poros la homogeneidad es un carácter que determina la pertenencia a un determinado grupo de los Boletales la parte himenial es heterogénea, o sea fácilmente separable.
En los Poliporales la parte himenial está unida de forma segura a la carne, por lo tanto no separable.
PIE
Si el carpóforo se presenta con sombrero y tallo es definido pedunculado; cuando el pie no está es definido sésil.
También para el tallo los caracteres a observar son muchos y todos revisten importancia fundamental para la determinación de la especie.
Forma del tallo y tipología del pie © Maria Tullii
Interior del tallo © Maria Tullii
Observaremos:
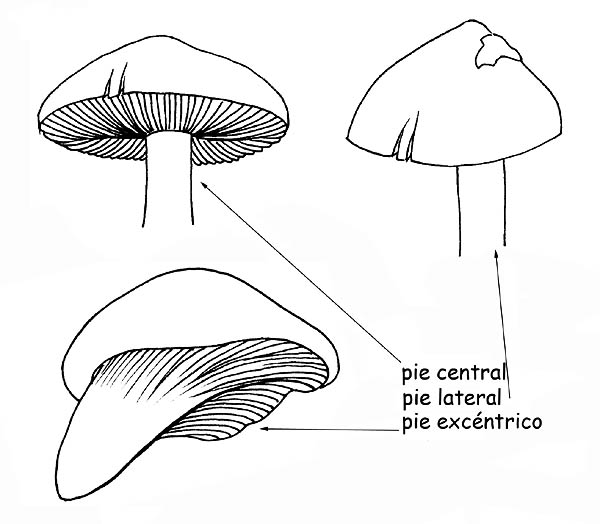
la inserción en el sombrero, puede ser central, lateral o excéntrica;
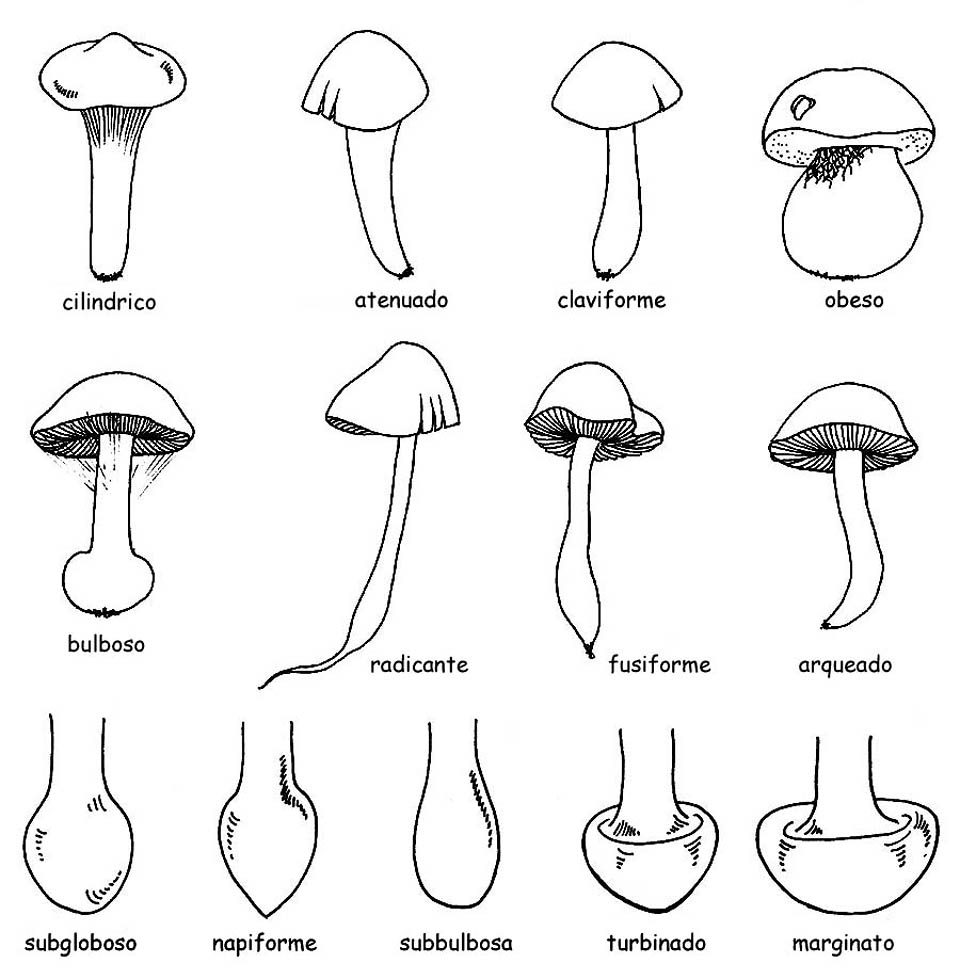
la forma: cilíndrica, atenuada hacia abajo o hacia arriba, obesa, ventruda o bulbosa.
Puede presentar un bulbo en la base: esto puede ser definido, según la forma, subesférico, napiforme, oval, turbinado y marginado.
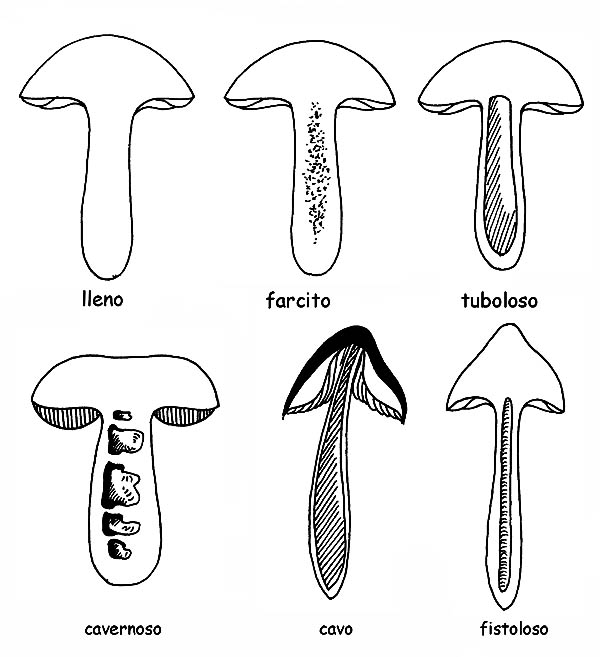
El interior puede presentarse carnoso, fibroso-carnoso, fibroso, lleno o hueco, fistuloso, cortical, etc.
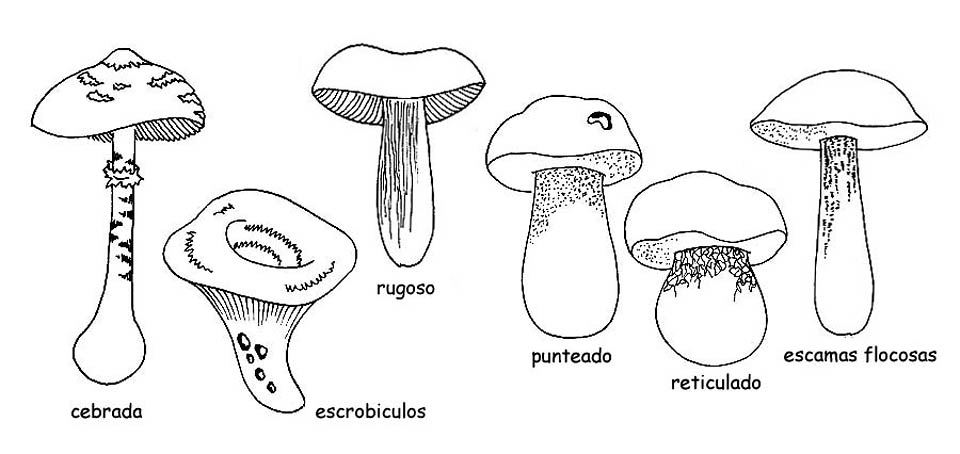
Pueden tener presente decoraciones de diversos tipos: por lo tanto la superficie puede presentarse lisa, algodonosa, escamosa, fibrilosa, verrugosa, peluda, punteada y enfundado, o decorada en modos diversos, pero también seca, viscosa o pegajosa.
Un discurso por separado, se debe hacer para las Boletaceae, donde la decoración del tallo están representadas mayormente por retículos. En este caso se debe observar el color, el tamaño de la malla de la red, si estas son ± alargadas o si son ± en relieve.
También el retículo puede estar ausente o el tallo puede presentarse recubierto por puntos o fibrillas, o también de asperezas o copos como en el género Leccinum.
VELOS
Todos los carpóforos en su estadio inicial de la vida (primocordio) están envueltos en una membrana cerrada que los envuelve (velo general).
En la mayor parte de los hongos este velo desaparece ya en los primeros días de desarrollo, pero en otros géneros, pertenecientes a los Agaricaceae, el velo generalmente persiste hasta la edad adulta y es comparable sobre el carpóforo bajo varias formas: verrugas o placas sobre el sombrero, volva o armilla en la base del tallo; siempre en las Agaricaceae, algunos géneros presentan otro velo: el velo parcial, que cubre la parte inferior del sombrero, partiendo del margen del sombrero y llegando hasta el tallo, protegiendo el himenóforo.
Decoraciones del tallo © Maria Tullii
Este velo primeramente cerrado en los ejemplares jóvenes, se depega del margen del sombrero a medida que el sombrero se abre, quedando pegado al tallo donde permanece, ± evidente, bajo la forma de anillo.
La volva puede ser membranosa, como una bolsa, envolvente, circuncisa, disociada en anillos o en perlas; el anillo puede ser algodonoso, largo, estrecho, ínfero (cuando se despega desde lo alto hacia abajo), súpero (cuando se despega desde abajo hacia lo alto) etc. si es residuo del velo general se llamará armilla. En algunas especies se encuentra presente un doble anillo, residuos del velo general y del velo parcial unidos entre ellos sobre el tallo; en otras el residuo del velo parcial está representado por filamentos sutilísimos y fugaces (cortina).
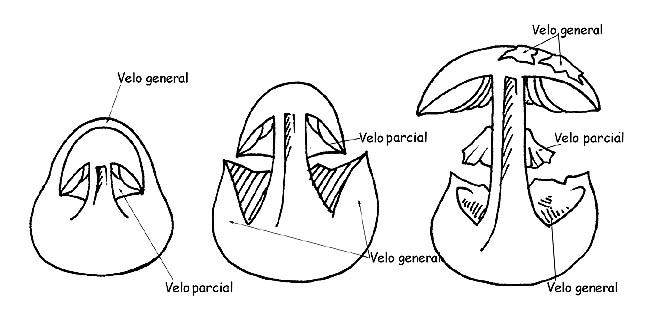
Desarrollo de los velos © Maria Tullii
CARNE
La pulpa de un hongo, o sea la parte que se observa cortando al hongo desde arriba hacia abajo se llama carne. Análogamente a las otras partes del hongo, también la carne presenta los caracteres fundamentales para la determinación de la especie: la consistencia puede ser sosa, compacta, tierna, coriácea, membranosa, cerosa, etc; el color y eventualmente la mutación del color de la carne expuesta al aire, debido a la oxidación de dos enzimas, carácter éste (particularmente importante para los Boletaceae) que viene observado con atención, anotando la intensidad y el tiempo de aparición del cambio de color. Puede tener tonalidades azules o rojas, según la dominancia de una enzima sobre la otra. La intensidad del cambio de color podrá ser mayor en los ejemplares jóvenes y frescos o más débil o inexistente en los ejemplares más viejos. La higrofaenidad es la capacidad ± evidente de la carne de absorber agua; la reviviscencia es la capacidad de la carne de desecar y volverse fresca en contacto con el agua; los caracteres organolépticos: olor y sabor de la carne son caracteres importantes, incluso determinantes en algunas especies del género Agaricus.
NOTAS DE SISTEMÄTICA
La micología es una rama de las Ciencias Naturales, aquel que trata de los hongos. Como cada ciencia, para ser clara, debe valerse de instrumentos de investigación y catalogación. Aquí está:
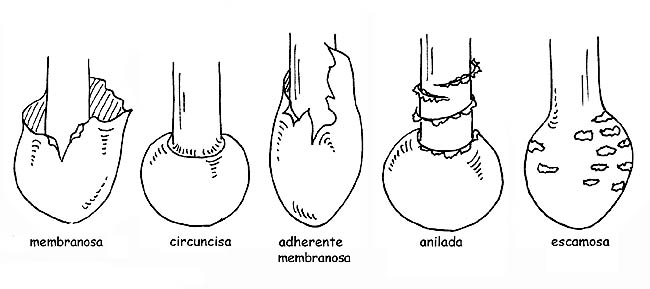
Tipologías de volva © Maria Tullii
la SISTEMÁTICA: es el instrumento científico que permite sistemar el objeto de estudio dentro de un esquema preciso (sistema) caracterizado por una subdivisión en compartimentos, cada uno con características comunes, en el interior de cada uno de los cuales están presentes otros compartimentos, continuar descendiendo hasta llegar a la unidad (una suerte de cajas chinas donde la especie es contenida en el interior de un género, el género en el interior de una familia, etc. mira los esquemas en la parte inferior). Es un instrumento en continua renovación y no unívoco, en cuanto puede ser interpretado de diversas maneras por diferentes micológos. La clasificación que proponemos es aquella que deriva de los padres de la sistemática moderna.
la TAXONOMÍA es la ciencia que estudia la clasificación del objeto de estudio, o sea el correcto posicionamiento de los varios taxa en el diseño sistemático. Por taxon se entiende una agrupación, de cualquier grado (especie, género, familia, orden, clase, división, reino).
la NOMENCLATURA, es el conjunto de reglas que consiente la asignación de un nombre científicamente válido y correcto al objeto de estudio. Estas reglas son sancionadas por el Código Internacional de Nomenclatura Botánica (C.I.d.N.B.). A este código hacen referencia todos los estudiosos y los autores de libros y publicaciones varias de botánica y micología. El C.I.d.N.B. ha sido fundado en 1867, cada 5-6 años se realizan Congresos y Comités de nivel mundial para aportar renovaciones y eventuales modificaciones.
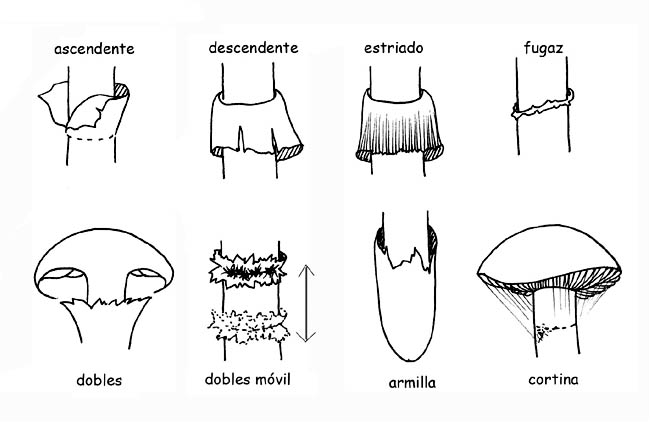
Tipologías de anillos © Maria Tullii
CLASIFICACIÓN Y ESQUEMA SISTEMÄTICO DE LOS HONGOS
Proponemos a continuación un esquema sistemático referido a la clasificación de los hongos, refiriéndonos a las enseñanzas derivadas de las escuelas de R. Kühner y R. Singer, famosos estudiosos de micología y según los dictámenes del C.I.d.N.B. (Código Internacional de Nomenclatura Botánica).
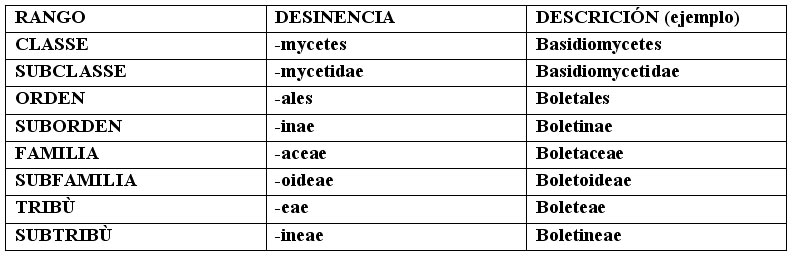
Nota: cuando se cita una especie, el nombre de esta se debe escribir en la forma binomial latina; el primer nombre, el del Género, debe tener la primera letra de la palabra en mayúscula (ej. Agaricus), el segundo, el de la especie, en minúscula (ej. campestris); sigue el nombre, normalmente abreviado, del autor que ha determinado y descripto al hongo (es. Bull. Por Bulliard); si (dentro de paréntesis), luego de este nombre hay otro, puede ser aquel del sancionador, y en este caso está siempre precedido de “:” y puede ser solo Persoon o Fries, (ej. Bull.:Fr.) o de quien ha completado la diagnosis original, en caso de ausencia del autor (por ejemplo la ausencia de la diagnosis latina), y en este caso el nombre viene precedido de “ex” (ej. Bull.ex Vent.). Puede luego estar presente un tercer nombre (fuera del paréntesis), que representa el autor que ha modificado la posición del Género o de la especie (ej. Bull.:Fr.)Kumm. La desinencia identifica los varios compartimentos de la sistemática. El Género, el subgénero, la sección, la especie, la variedad y la forma tienen desinencia variable, ligadas a la raíz lingüística. Hacen excepción Reino y División, que no tienen particulares desinencias. De la Clase a la Subtribu, en orden descendente:
MICROSCOPÍA
Como se ha visto para los caracteres macroscópicos, también los caracteres microscópicos de los hongos revisten importancia fundamental para la correcta identificación de muchas especies. (por ej. En el Género Mycena). En esta, que quiere ser una disertación divulgativa y no estrictamente científica, ilustraremos las principales estructuras microscópicas presente en los hongos superiores, los cuales se dividen en dos grandes clases: Ascomycetes y Basidiomycetes. Ilustraremos en breve también los reagentes y las técnicas más comunmente usadas para la observación.
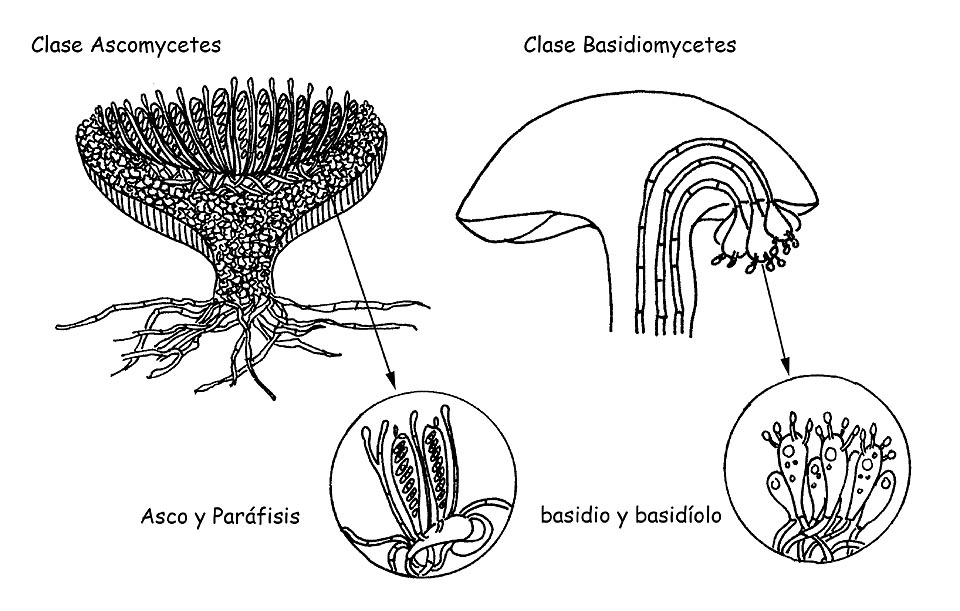
Hongos superiores © Maria Tullii
ASCOMYCETES: SUS ELEMENTOS MIROSCÓPICOS
Ascomicetos © Maria Tullii
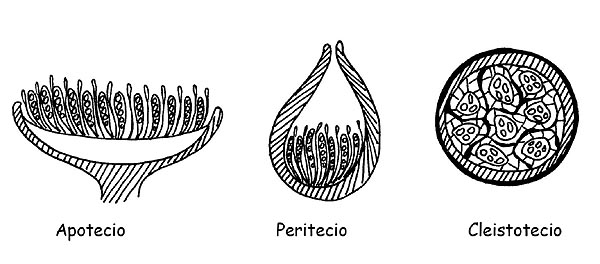
Es importante, para esta clase de carpóforos, es conveniente realizar una indicación sobre el desarrollo de la parte fértil de hongo: el himenio (tecio).
Cuando el carpóforo expone completamente al aire su parte himenial diremos que estamos en presencia de un Apotecio, cuando la parte himenial comunica con el exterior solo a través de una pequeña abertura apical diremos que se trata de un Peritecio y cuando finalmente no está en contacto con el exterior diremos que se trata de un Cleistotecio (en este caso la mayor parte de la especie tienen desarrollo hipogeo o semi-hipogeo).
Veremos que esta distinción es importante desde un punto de vista microscópico para la observación específica de los varios elementos.
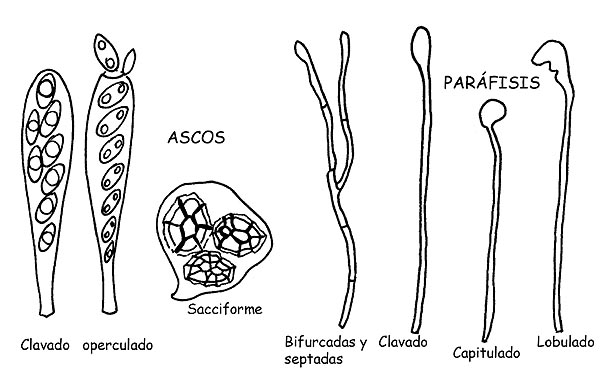
Ascos y paráfisis © Maria Tullii
Los ascos, de donde deriva la denominación de la clase, son los elementos fértiles que producen las esporas y se desarrollan de particulares hifas especializadas, llamadas ascogenas.
Se presentan cilíndricas ± claviformes o en forma de odre o se saco y contienen en su interior las esporas (mayormente en número de ocho, pero también en número variable, hasta treinta y dos o más) hasta la maduración. Pueden tener pared simple o doble (tunicada), pueden tener reacciones positivas o negativas al reactivo de Melzer, en caso de tener reacción positiva se dice amiloide o yodos positivos.
Los ascos cilindroides, típicos de los Apotecio y de los Peritecio, se abren con la maduración de las esporas, por laceración o grieta de la parte apical o mediante la apertura de un minúsculo opérculo dejando salir las esporas maduras.
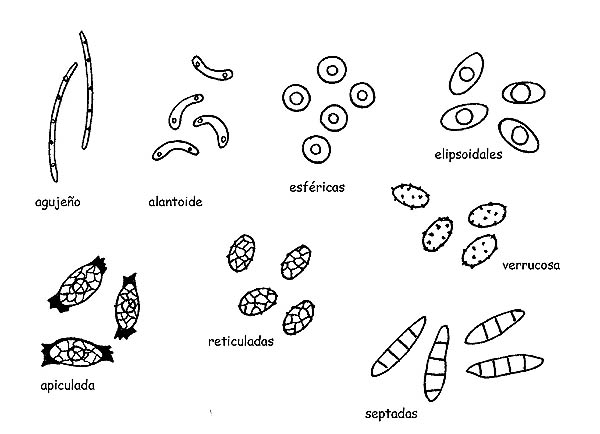
Esporas © Maria Tullii
Los ascos sacciformes típicos en llugar de los Cleistotecios, liberan las esporas por la descomposición de las paredes.
Las Paráfisis son sutiles hifas estériles, mixtas en las ascas y cumplen una función de sostén para estos últimos.
Se presenta mayormente de forma cilíndrica, pueden ser bifurcada o septada, y el ápice se puede presentar clavado, lanceolado, capitulado, lobado, incrustado, etc.
También en la parálisis, para la identificación de algunas especies, se observa la positividad al reactivo de Melzer.
Las Esporas en los ascomycetes asumen una gran variedad de formas (acicular, alantoide, esférica, elipsoidal, apiculada, etc.) colores y ornamentaciones (retículos, crestas, verrugas, espiga, etc.) pueden ser aserradas septadas ya sea longitudinalmente como transversalmente.
Los Pelos son elementos diferenciales para muchas especies de ascomicetes, protegen el himenóforo y retienen la humedad necesaria para el desarrollo de la especie.
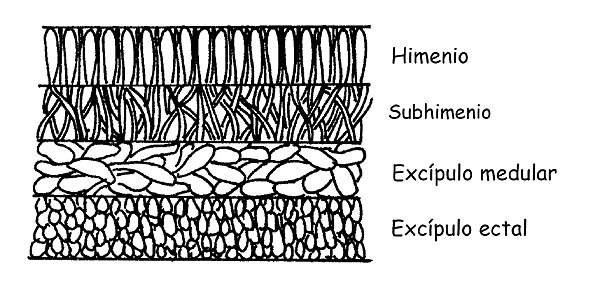
Sección del apotecio © Maria Tullii
Pueden estar desplazados sobre la superficie externa o sobre el margen del carpóforo, presentan una gran variabilidad de formas y dimensiones, pueden ser septados o no, con ápice agudo o ± redondeados, etc.
La Textura: por textura se entiende la esctructura de la carne que puede ser formada por células de diversa forma y disposición.
Es además importante en los Apoteci, observar la sección completa del carpóforo.
Esta está constituida por más estratos: Himenio, Subhimenio, Excípulo medular, Excípulo ectal.
La observación de la conformación de las hifas de los simples estratos puede ser determinante para llegar a la determinación de una especie.
BASIDIOMYCETES: SUS ELEMENTOS MICROSCÓPICOS
En los Ascomycetes hemos visto que las esporas se desarrollan en el interior de estructuras especializadas, los asca, que la protegen hasta la completa maduración. Al contrario, los Basidiomycetes desarrollan e maduran sus esporas en el exterior de estructuras fértiles, los Basidios.
La clase Basidiomycetes se divide en dos grandes subclases: Holobasidiomycetes e Heterobasidiomycetes. Pertenecen a la primera todas las especies con basidios monocelulares y a la segunda con basidios pluricelulares.
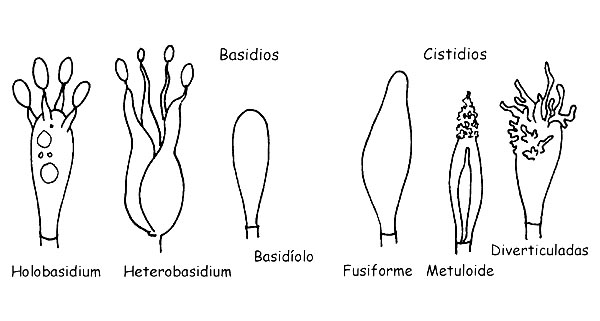
Estructura de los Basidiomycetes © Maria Tulliii
Los Basidios son células especializadas por la producción de las esporas. Nacen de la célula terminal de una hifa. En el interior del basidio se realiza la división celular que da origen a cuatro células cada una de las cuales se transformaré en una espora.
La formación de las esporas, sucede en la cima de las protuberancias puestas en el ápice del basidio (esterigma) y, en la maduración, las esporas vendrán relanzadas en el ambiente. Los basidios son generalmente ± cilindricos y ± clavados, presentan generalmente cuatro esterigmas (basidios tetrasporicos) pero pueden también presentar dos o tres o más de cuatro, pueden o no presentar pigmento celular.
Los Basidiolos son basidios en formación
Los Cistidios son elementos estériles y pueden estar presentes ya sea en la parte himenial como sobre la cutícula o sobre el tallo. Los cistidios presentes en las láminas se llaman: Pleurocistidos si están presente en los rostros laminar, Cheilocistidios si están presentes sobre en fila de la lámina, mientras que aquellos de la cutícula y del tallo se llaman respectivamente Dermatocistidios y Caulocistidios. Los cistidios pueden presentarse en formas muy diferenciadas de especie en especie, con o sin incrustaciones, con paredes simples o dobles, con o sin pigmento intracelular. Revisten por lo tanto gran importancia desde un punto de vista microscópico en cuanto consienten a menudo una correcta determinación.
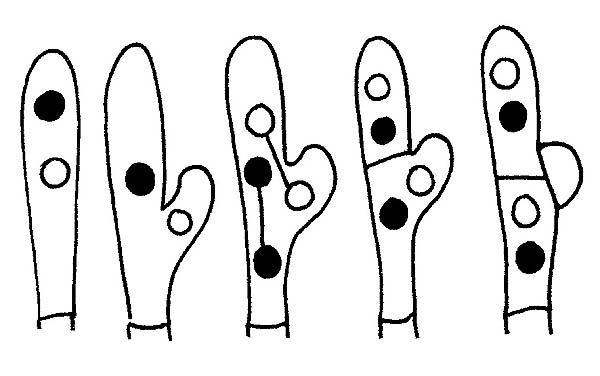
Formación de las fíbulas © Maria Tullii
Las Esporas también en los basidiomycetes se presentan muy variables, en la forma, en el color y en las ornamentaciones. Presentan generalmente una pequeña protuberancia en una de las dos extremidades (apiculo) que no es otro que el apéndice que tenía anclada la espora en el esterigma, y pueden tener en la otra extremidad un poro germinativo, una suerte de florcita de donde tendrá origen el nuevo micelio. También por las esporas de los basidiomycetes un carácter importante por la determinación es la amiloidea o sea la reacción positiva al reactivo de Melzer, la cianofilia es decir cuando la pared esporal se tiñe de azul en contacto con el reactivo Azul algodón.
Las Fíbulas son el resultado de un particular tipo de crecimiento de las hifas mediante un divertículo producido por las células apicales. No están presentes en todos los géneros o en todas las especies de un género y su presencia o ausencia es a menudo decisiva por la determinación de la especie en examen. Se encuentran generalmente en los septos de las hifas pero también en la base de los basidios o basidioles. En los baisdiomycetes reviste particular importancia, sobre todo para algunos géneros, el estudio de la forma y de la disposición de las hifas que constituyen la Pileipellis (revestimiento del sombrero), particularmente importante por ejemplo en el género Russula, y la Caulopellis (revestimiento del tallo), por ejemplo en el género Mycena.
PRINCIPALES REACCIONES Y MÉTODOS DE PREPARACIÓN DE PORTAOBJETOS DE MICROSCOPIO
AMILOIDIA
sobre muestra seca:
1) rehidratar el fragmento a observar en KOH 5% por cerca 1’
2) hacer uno o dos lavados en agua y secar.
3) inserir una gotita de Melzer y esperar 1’ ca.
4) lavar con Cloralio Idrato y secar
5) observar en agua o en Cloralio Idrato o bien en L4 Tampon a la glicerina
sobre material fresco:
1) excluir el pasaje en KOH 5%
2) proceder como el punto 3
CIANOFILIA
sobre muestra seca:
1) re hidratar por cerca 1’ en KOH 5%
2) lavar y secar todo por 1-2 veces en agua
3) inserir una gota de Blu lactico
4) calentar el preparado sobre fuego moderado por algunos segundos, si es necesario agregar mas Blu láctico
5) quitar el fragmento y pasarlo sobre otro vidrio portaobjeto
6) agregar Cloral hidratado
7) observar
NOTA: para algunos géneros como Galerina, Rhodocybe o Lepista no es necesario calentar el preparado.
CARMINOFILIA O SIDEROFILIA
sobre muestra seca:
1) rehidratar el fragmento en KOH 5% por cerca 1’
2) lavar y secar el preparado por 1-2 veces en agua.
3) poner Carmín Acético.
4) calentar todo a fuego muy lento agitando con un hierro (no acero) por algunos segundos.
5) pasar el preparado a otro vidrio portaobjeto y agregar Carmín Acético o Cloral Hidratado.
6) apretar muy fuerte el preparado y observar.
PRINCIPALES REAGENTES Y PREPARADOS PARA LA OBSERVACIÓN MICROSCÓPICA
Mesa de trabajo © Pierluigi Angeli
AMONIACO (NH3) : se utiliza para la rehidratación de las muestras secas (observación de muestras secas) ; normalmente está en solución acuosa concentrada al 2-3 o 6%.
HIDRATO DE POTASIO (KOH) : tiene las mismas propiedades que el amoníaco, se usa por lo tanto en muestras secas en una concentración máxima del 5%.
HIDRATO DE SODIO (NaOH) : idéntico a los precedentes, también éste se usa en una concentración no superior al 5-10%.
HIDRATO DE CLORAL (C Cl3 CH(OH)2) : tiene la característica de aclarar.
En el caso de que se quiera observar por mucho tiempo el preparado es necesario usar líquidos que eliminan la posibilidad de reproducción de los microbios en el interior del vidrio portaobjeto, el LACTOFENOL DE AMMAN (solución de Amman) compuesto por Fenol, Ácido láctico y glicerina en solución acuosa al 20 % es un líquido bactericida muy indicado, especialmente si se quieren hacer largas observaciones siguiendo fotomicrográficas; es importante fijar bien el vidrio cubreobjetos al portaobjetos,
se obtiene poniendo sobre el vidrio cubreobjeto un pequeño peso y sellando los bordes con esmalte para uñas) impidiendo eventuales escapes de líquido o infiltración de aire.
Las soluciones amoniacales y la potasa se usan también para la observación de material fresco, en efecto en algunos géneros (Galerina, Inocybe, Cortinarius) colorean las paredes de las células marginales (cistico) provocando una reacción importante desde el punto de vista taxonómico.
ROJO CONGO: es el colorante más usado en Micología, colorea las paredes de las células, se adapta bien en muchas estructuras celulares; si es usado en solución amoniacal es óptimo también para trabajar con muestras secas.
FLOXINA: Tiene las mismas características que el Rojo Congo, colorea el citoplasma, es mayormente usada por los micólogos americanos, también ella se encuentra en solución acuosa como en solución amoniacal.
AZUL ALGODÓN O AZUL DE METILENO: el más apto es el Azul Algodón en Lactofenol, se usa también como reagente para individualiza la cianofilia esporal, o sea la membrana esporal asume una coloración intensa (género Entoloma) o si no se colorea (género Tricholoma).
EAGENTE DE MELZER: Se obtiene mezclando una solución de Iodo Cloral Hidratado; sirve principalmente para verificar la amiloidea de las esporas. La reacción puede ser amiloide (membrana esporal gris-negruzca o azulada, ápice de los ascos verde-azul) destrinoide o pseudos amiloide (la espora asume una coloración rojiza), inamiloide.
CARMIN ACÉTICO: se usa para observa una particular reacción que sucede en el interno de los basidios de las especies pertenecientes al grupo del Lyophyllum s.l. Las granulacions de estos basidios se cefinen como carminófilas o siderófilas y asumen una coloración téipicamente gris-negruzca.
NEO-FUCSINA BASICA (reactivo de ZIEHL) : produce una reacción sobre las incrustaciones hifales en la cutícula de algunas Russula, asumiendo un importante valor taxonómico.
BLU CRESILE : produce una reacción llamada metacromática en las hifas de algunas especies del género Mycena. La reacción se manifiesta transformando la pared hifal externa en un vivaz rojo-violáceo.
SULFOVAINILLINA : pone en evidencia las hifas laticífera y los cistidios en los géneros Russula y Lactarius.
SULFATO FERROSO (FeSO4) : se utiliza como reactivo macroquímico en el género Russula.
TINTURA DI GUAYACO : también este es un reactivo muy usado en el género Russula.
MEDICIÓN: QUE. COMO Y CUANTO
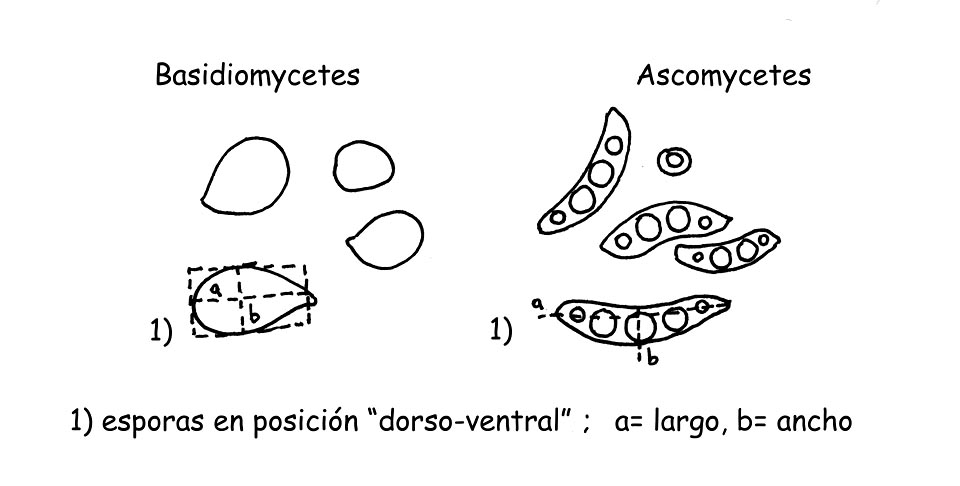
Medición esporas © Maria Tullii
Esporas Para la medición de las esporas se usa normalmente relevar el lorgo y el ancho, imaginando una hipotética forma geométrica (un cuadrado, un rectángulo o un círculo), excluyendo las ornamentaciones y el apéndice hilar. Deben ser medidas las esporas cuando se presentan en la posición “dorsoventral”, como la figura (A), normalmente se efectúan cerca de 32 mediciones de esporas de carpóforos maduros, mejor sería efectuar la medición sobre esporada. Para una correcta y válida exposición de las medidas efectuadas es necesario calcular ya se los “Q” y “QM” (o sea el cociente medio) sea el Volumen “V” y volumen medio “VM” de las esporas.
Para el “QM”:
se eliminan las mediciones marcadamente anómalas, ya sea por aquellas muy pequeñas o para aquellas muy grandes, luego de cada medición se encontrará el cociente, (es. 10 × 6,5 – 10,5 × 7 – 9,5 × 7 etc. 10/6,5 = 1,53846153 que se redondeará a 1,53 + 1,5 + 1,35 ecc. = 4,38, dividiendo luego por el número de las mediciones del cual ha sido encontrado el cociente se obtendrá el “QM”, en este ejemplo 4,38/3 = QM 1,46;
Para el “Q”:
se reportará, de los cocientes obtenidos para el “QM” el más chico y el más grande como para indicar la medición de las esporas Q= 1,35 – 1,53.
En cuanto al volumen de las esporas se continúa así:
se relevará la espora más pequeña de nuestro hongo, de ella se calculará el volumen con la siguiente fórmula V= 4π/3 × (a/2)² × b/2 (a = diámetro más pequeño, b = diámetro
más grande).
Se relevará la espora más grande y se calculará el volumen como para la precedente V1= 4π/3 × (A/2)² × B/2 (A = diámetro más pequeño B = diámetro más grande) se proseguirá luego con la fórmula VM = (V + V1)/2, por ejemplo: en una serie de mediciones la espora más pequeña que mide 6,5 × 4 µm y la más grande 8 × 5,5 µm aplicando la fórmula tendremos: V = (4×3,14/3) × (4/2)² × 6,5/2 = 54,34; V1 = (4×3,14/3) × (5,5/2)² × 8/2 = 126,40; VM = (54,34 + 126,40) / 2 = VM 90,4.
Basidios y Cistidios : se efectúa la medición del largo, partiendo de la unión del basidio o del cistidio al bajo himenio, el ancho se toma en el punto medio o normalmente en el punto más ancho.

Amanita muscaria – Amanita phalloides – Astraeus hygrometricus

Astraeus hygrometricus – Boletus aestivalis – Boletus edulis – Suillus granulatus

Tricholoma columbetta – Suillus granulatus – Tuber melanosporum – Tuber magnatum – Tuber melanosporum – Macrolepiota procera

Macrolepiota procera – Russula vesca – Otidea onotica

Verpa bohemica – Morchella esculenta – Marasmius oreades – Mutinus elegans – Macrolepiota procera

Leucoagaricus americanus – Amanita citrina – Amanita muscaria

Amanita muscaria – Amanita rubescens – Boletus aereus

Boletus aereus – Laccaria amethystina – Cortinarius rufoolivaceus

Cortinarius rufoolivaceus – Gyromitra infula – Cortinarius lividoviolaceus

Cortinarius lividoviolaceus – Amanita spissa


Amanita spissa – Cantharellus cibarius – Amanita mairei

Amanita mairei – Clitocybe gibba

Calvatia excipuliformis– Calvatia craniiformis – Clavariadelphus pistillaris

Clavariadelphus pistillaris – Clitocybe nebularis

Cortinarius turmalis – Clavulina cinerea – Gomphidius glutinosus

Gomphidius glutinosus – Clitocybe odora

Russula atropurpurea – Calvatia utriformis – Crucibulum laeve

Lactarius deliciosus – Suillus grevillei

Suillus grevillei – Russula mairei – Leccinum aurantiacum

Leccinum aurantiacum – Mycena inclinata – Oudemansiella mucida

Oudemansiella mucida – Mycena renatii – Lepiota clypeolaria – Lactarius torminosus

Lactarius torminosus – Badhamia utricularis

Badhamia utricularis

Clavulina rugosa

Russula foetens – Psathyrella caput-medusae – Cantharellus lutescens – Suillus luteus

Suillus luteus – Amanita muscaria – Stropharia aeruginosa – Tricholoma saponaceum


Amanita pantherina – Aleuria aurantia

Boletus aestivalis – Entoloma lividoalbum

Boletus edulis – Boletus aestivalis

Chroogomphus rutilus – Calocera viscosa – Gloeophyllum sepiarium

Stereum hirsutum – Lactarius aurantiacus – Fuligo septica

Fuligo septica

Fuligo septica – Cortinarius variecolor – Fistulina hepatica

Fistulina hepatica – Galerina marginata – Hypholoma sublateritium

Hypholoma sublateritium – Hygrophoropsis aurantiaca – Hypholoma fasciculare

Hypholoma fasciculare – Ischnoderma benzoinum

Ischnoderma benzoinum – Leucocortinarius bulbiger – Leccinum holopus – Spathularia flavida

Spathularia flavida – Paxillus involutus

Clitocybe costata – Fomitopsis pinicola – Lycoperdon perlatum

Lycoperdon perlatum – Lepista flaccida = Lepista inversa – Suillus grevillei – Russula integra

Russula integra – Lactarius intermedius – Oudemansiella radicata

Oudemansiella radicata – Suillus granulatus – Clathrus ruber

Clathrus ruber – Tricholoma acerbum – Russula xerampelina

Ramaria lacteobrunnescens – Armillaria mellea = Armillariella mellea


Amanita vaginata – Amanita phalloides

Clitocybe costata – Boletus aestivalis – Boletus regius


Boletus edulis – Agaricus silvicola – Clitocybe candicans – Amanita rubescens

Cantharellus cibarius – Chroogomphus rutilus

Calocera viscosa – Clavulina rugosa

Cortinarius anomalus – Cortinarius cliducus – Cortinarius infractus

Cortinarius olivascentium – Cystolepiota seminuda

Cribraria argillacea – Ceratiomyxa fruticulosa

Ceratiomyxa fruticulosa = Famintzia fruticulosa

Ceratiomyxa fruticulosa var. porioides

Clavulina coralloides

Lactarius piperatus – Coprinus micaceus

Collybia fusipes – Cortinarius cotoneus – Dacrymyces abietinum

Ife miceliari

Ife miceliari – Hemimycena gracilis – Hemimycena pseudolactea


Hypholoma fasciculare – Hebeloma edurum

Lycoperdon perlatum – Lactarius deliciosus

Lactarius salmonicolor – Lactarius volemus

Lactarius porninsis – Lactarius serifluus – Leccinum aurantiacum

Lycogala epidendron

Lycogala epidendron

Lactarius circellatus – Lactarius intermedius – Laetiporus sulphureus – Hypholoma sublateritium – Leccinum carpini

Leucopaxillus candidus – Macrolepiota mastoidea

Lycoperdon echinatum – Lycoperdon molle

Macrolepiota procera – Mycena pura

Phaeomarasmius erinaceus – Merulius tremellosus

Phellodon tomentosus – Pluteus cervinus

Polyporus brumalis – Paxillus involutus

Russula cyanoxantha – Russula risigallina

Russula virescens – Russula aurea

Russula integra – Russula velutipes = Russula aurora

Russula delica – Marasmius oreades – Marasmius bulliardii – Scleroderma citrinum

Sowerbyella imperialis

Stropharia caerulea – Suillus granulatus – Trametes versicolor – Penicillum digitatum

Stemonitis axifera : fase juvenil

Stemonitis axifera : juvenil, en crecimiento y adulta

Xerocomus subtomentosus – Xerocomus chrysenteron – Leccinum scabrum

Amanita rubescens – Amanita lividopallescens
Boletus satanas

Boletus satanas – Pholiota squarrosa
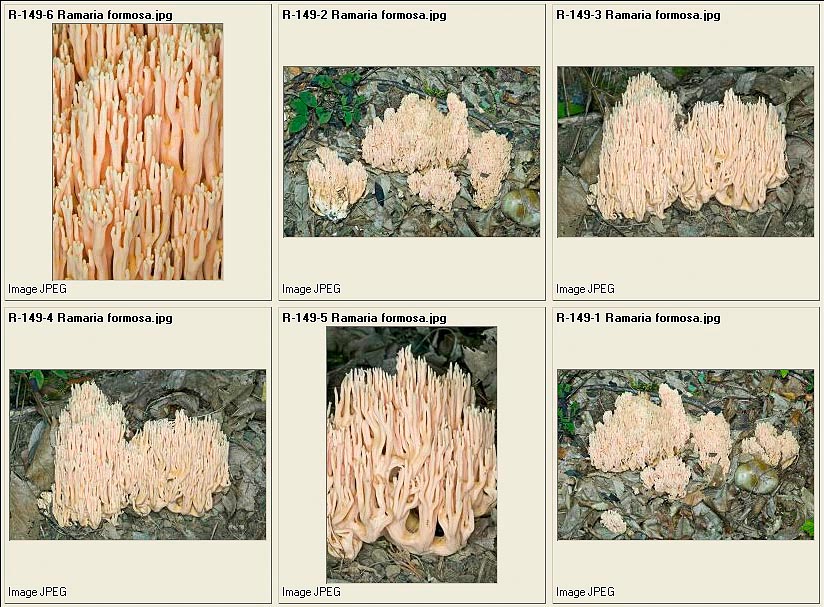
Ramaria formosa
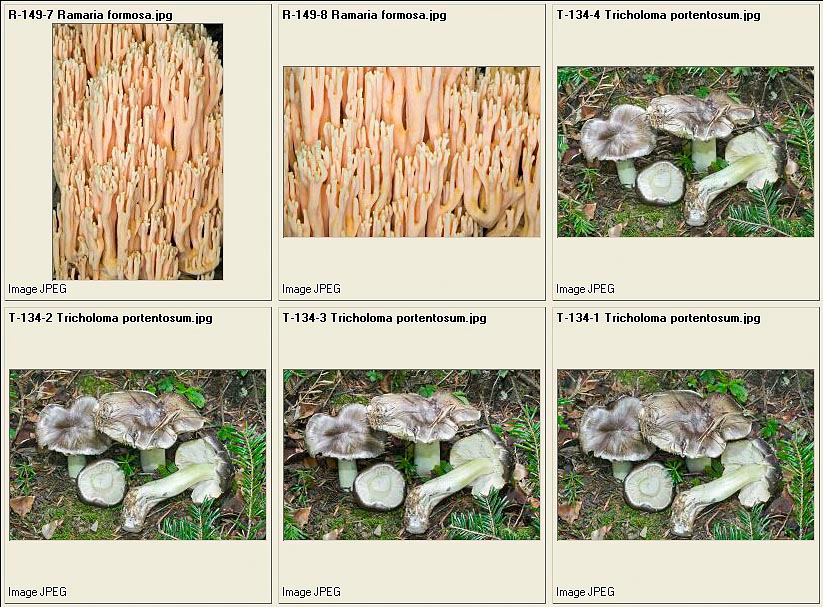
Ramaria formosa – Tricholoma portentosum

Boletus edulis en edad madura – Crucibulum laeve

Lasiochlaena benzoina = Ischnoderma benzoinum – Lycoperdon echinatum

Trametes versicolor – Trametes gibbosa

Phaeolus schweitnizii – Coprinus niveus

Cortinarius herculeus – Kretzschmaria deusta = Ustulina deusta : la forma gris es el Anamorfo, la negra es el Teleomorfo

Inocybe geophylla – Tricholoma terreum – Perenniporia narymica – Laccaria bicolor

Stropharia caerulea – Coprinus comatus

Coprinus comatus – Amanita caesarea – Spore di Tuber melanosporum

Esporas de Boletus edulis – Otidea umbrina – Tuber magnatum
© Giuseppe Mazza