

di Pierluigi Angeli e Maria Tullii
BREVI CENNI DELLA STORIA DELLA MICOLOGIA

Agaricus praeclaresquamosus © Giuseppe Mazza
MICOLOGIA, la scienza che studia i funghi, viene dal greco MÝCHÊS = fungo e LÒGOS = discorso, trattato. Vuol dire dunque letteralmente trattare di funghi, e micologo è colui che studia i funghi.
Per parlare di funghi e capire certi atteggiamenti e usanze, è utile, a volte, partire anche da molto lontano: dalla preistoria.
In base alle testimonianze rinvenute (graffiti) si può supporre che l’uomo abbia osservato i funghi fin dai tempi più remoti, e abbia imparato ad utilizzarli in primis come cibo.
Testandone la pericolosità sulla propria pelle, ha poi scoperto le proprietà particolari di alcune specie: i funghi allucinogeni, usati da stregoni e sciamani, e quelli curativi, dotati di capacità antinfiammatorie o rafforzanti del sistema immunitario.
In tempi relativamente più recenti i funghi mortali sono stati poi spesso usati per eliminare nemici e personaggi scomodi.
Come si può molto facilmente immaginare all’inizio gli avvelenamenti da funghi erano molto frequenti e la difficoltà di capirne le cause fece si che nascessero superstizioni e false credenze, che ancora oggi sopravvivono in molti luoghi. Si pensava che i funghi, innocui per loro natura, diventassero tossici a causa di fattori esterni, oppure si vedeva in essi una creazione del diavolo per attirare l’uomo e poi nuocergli.

Amanita pantherina © Giuseppe Mazza
Questa concezione del fungo può essere ricondotta al modo di nascita e crescita del fungo stesso: esso sorge dalla terra apparentemente dal nulla (la “pianta” fungo si sviluppa sotto terra, non produce “semi”, nell’accezione comune del termine, e la crescita dei carpofori non è strettamente legata ad un ciclo stagionale preciso, seguendo più le condizioni climatiche, variabili di anno in anno); inoltre la crescita dei carpofori si sviluppa spesso in cerchi o linee a zigzag, che venivano scambiati per disegni del maligno.
La prova che alcune specie erano usate per provocare allucinazioni e stati di trance in riti sciamanici o divinatori, la troviamo nei molti graffiti e disegni rinvenuti in varie parti del mondo: nel deserto del Sahara, nel territorio di Algeri, nella Valle delle Meraviglie in Francia, nello stato Indiano, nel nord dell’Europa; ovunque, le raffigurazioni rappresentano riti di gruppo e i funghi rappresentati fanno pensare al genere Psilocybe o comunque a funghi sicuramente con potere allucinogeno.
Quello divinatorio non era però l’unico uso che si faceva dei funghi: ne abbiamo una testimonianza recente. Nel 1991 sul ghiacciaio del Similaun, che divide l’Italia dall’Austria, è stato ritrovato il corpo mummificato di un uomo morto oltre 5000 anni fa. Tra gli oggetti che facevano parte del suo corredo, c’erano anche il Fomes fomentarius e il Lenzites betulinus; il primo serviva per accendere il fuoco, il secondo era usato in decotto per aumentare la resistenza fisica alla fatica e al dolore, in polvere come antibiotico ed emostatico per curare le ferite e anche, probabilmente, come spazzola per detergere la pelle.

Il Fomes fomentarius serviva nell’antichità per accendere il fuoco © Pierluigi Angeli
Tutt’ora, nei paesi scandinavi, il Lenzites betulinus, considerato un corroborante naturale, viene consumato in decotto. In tempi più recenti si cominciano a trovare, anche se non in quantità, testimonianze scritte sui funghi, la loro natura, il loro utilizzo, le credenze ad essi legate. Uno dei primi scritti documentati sui funghi, in un’opera dell’antica Grecia, parla di come si coltivano: “… interrare del letame equino sotto un albero di fico ed innaffiarlo abbondantemente …”.
Il greco TEOFRASTO (370-287 a.C.) nativo dell’isola di Lesbo e discepolo di ARISTOTELE, che è considerato il padre della botanica, parla di funghi così: “… essi sono piante imperfette, prive di radici, di foglie, di fiori e di frutti”.
Dopo TEOFRASTO la materia funghi passò nella sfera di competenza dei medici e bisogna arrivare al 50 d.C. per trovare ancora scritti sui funghi.
Nell’opera “DELLA MATERIA MEDICA” di PEDANO DIOSCORIDE, nato in Cilicia (Asia Minore), medico militare e civile in Roma sotto gli imperatori Claudio e Nerone, egli ci ha tramandato alcune nozioni sulla tossicità dei funghi e sulle terapia da seguire in caso di avvelenamento: “… decotti di erbe aromatiche, pozioni di aceto e sale, somministrazioni di sterco di pollo impastato con il miele e aceto …”, rimedi che probabilmente sono stati usati per molti secoli. A lui risale la prima descrizione dell’AGARICUM (Laricifomes officinalis) che era usato come medicinale.

Boletus satanas © Giuseppe Mazza
Un altro medico che si occupò di funghi, greco di Pergamo che visse a Roma al tempo degli imperatori Marco Aurelio, Lucio Vero e Commodo fu CLAUDIO GALENO (129-200 d.C.). Galeno aveva suddiviso i funghi in tre gruppi fondamentali: “Bolites”, “Amanitai” e “Mikés”.
Al primo gruppo appartenevano i funghi a lamelle (tra cui le attuali Amanite, genere al quale appartiene Amanita caesarea che ancora oggi, in alcuni luoghi d’Italia, viene comunemente chiamata “boleto”); al secondo gruppo tutti i funghi a tubuli e pori (l’attuale genere Boletus, secondo la nomenclatura creata da Linneo nel 1735); al terzo gruppo infine facevano capo tutti gli altri funghi a cappello e gambo. Secondo Galeno i funghi appartenenti ai primi due gruppi erano da considerare tutti commestibili, mentre quelli del terzo gruppo erano da considerare tossici e non adatti al consumo perché: “… non nutrienti, difficili da digerire e pericolosi per le proprietà tossiche …”. Galeno confermava infine la validità della terapia a base di sterco di pollo contro gli avvelenamenti.
Il primo autore che trattò i funghi in modo più approfondito, è Gaio Plinio Secondo detto PLINIO IL VECCHIO (23-79 d.C.). Egli ci ha lasciato un’opera in 37 libri: “HISTORIA NATURALIS” dove, fra gli altri argomenti naturalistici, parla anche di funghi. Eccone alcuni stralci: “ … fra i cibi meno raccomandabili crediamo si debbano annoverare i boleti …” (le attuali Amanite ndr) e motiva questa sua affermazione con la facilità di scambio con funghi tossici.

Amanita muscaria © Giuseppe Mazza
Riferendosi all’ Amanita muscaria dice: “ … alcuni si distinguono per una sorta di verruche che sono sul cappello, bianche e secche, simili ad efflorescenza di salnitro, e che altro non sono che i residui del velo …”.
Inoltre cita alcune circostanze in cui funghi perfettamente eduli possano diventare tossici e dice: “ … se nascono in vicinanza di bottoni metallici, chiodi da scarpa, ferri arrugginiti, panni fradici, assorbendo i succhi impregnati di tali sostanze e trasformandoli in veleno”.
E ancora: “ … se nelle vicinanze vi è qualche tana di serpente, o se un serpente nel suo passaggio vi soffia sopra, il fungo diventa velenoso, perché la sua natura è di assorbire qualunque tipo di sostanza velenosa”.
Queste credenze sono ancora fortemente presenti e ritenute valide in quasi tutto il territorio italiano.
Si credeva anche che la commestibilità dei funghi si poteva intuire osservando le piante presso cui crescevano. Plinio definiva a questo proposito innocui quelli che nascono sotto le conifere, il fico e la ferula; tossici quelli che nascono sotto le querce, i faggi e i cipressi.

Amanita phalloides © Giuseppe Mazza
Questo probabilmente perché le Amanita mortali e la phalloides in particolare prediligono le latifoglia, mentre sappiamo che sulle radici della ferula vi nasce il Pleurotus eryngii, ottimo fungo conosciuto fin dalla antichità.
Plinio descrive anche lo sviluppo dei boleti: “… nascono dalla terra, dapprima tutti avvolti nella loro volva, che li fa assomigliare a uova. A tale stadio di sviluppo questi funghi sono particolarmente apprezzati. In seguito la volva, rompendosi, rimane attaccata alla base del gambo …”.
Per la prima volta viene usato il termine volva nel suo corretto significato micologico. Egli descrive poi la durata di un boleto che è di soli sette giorni, la crescita non cespitosa, con carpofori non appaiati ma singoli.
Per spiegare la natura dei funghi (che nascono apparentemente dal nulla) Plinio dice: “… la loro origine va ricercata nel limo della terra umida e nei suoi umori, che incominciano a fermentare, oppure nelle radici delle piante cupulifere …”.
Questa affermazione restò radicata per secoli nelle opinioni dei naturalisti, fino a quando, con le nuove strumentazioni, si riuscì a dare una connotazione più scientifica allo studio della materia. Del boleto Plinio ne fa un fungo nobile e lo tratta a parte, mentre relega tutti gli altri insieme e dice: “ … altri funghi hanno una consistenza più soda. Di essi esistono infiniti generi, ma hanno tutti una comune origine: la linfa degli alberi. Sono commestibili sicurissimi quelli di colore rosso più cupo dei boleti …”.
In questo scritto parla, con ogni probabilità, di funghi lignicoli e senz’altro di Fistulina epatica, che era considerata ottimo commestibile fin dai tempi antichissimi.

Fistulina hepatica © Giuseppe Mazza
Dice ancora: “ … vi sono poi specie di colore bianco, con gambo molto sviluppato e cappello che assomiglia al copricapo di un Flamine …”. Il Flamine era un sacerdote di una antica religione Romana che aveva un copricapo che nell’aspetto ricordava una Macrolepiota procera con cappello non ancora del tutto aperto.
E: “ … un terzo genere di funghi, i Suilli è molto incline al veleno …”.
I Suilli erano le attuali Boletaceae, ed erano considerate velenose per il colore che diventa livido, ossia bluastro. Per contro, al tempo di Plinio i “Suilli” erano un prodotto molto commercializzato: venivano importati dall’Asia Minore e confezionati infilzati in un giunco sottile e messi a seccare vicino al fuoco, sistema usato ancora oggi in alcuni luoghi del nostro paese. Inoltre, alcuni Suilli erano usati dai medici per curare emorroidi, brufoli, morsicature ed anche per preparare un collirio.
Ma il pericolo maggiore, affermava Plinio, veniva dai porcini tossici che hanno fatto strage di intere famiglie. Plinio, nel suo trattato, consiglia: “ … se qualcuno vuole proprio cibarsene cerchi di fare attenzione al momento dell’acquisto e curi particolarmente la loro cottura …” E conclude: “ … si diffidi di quei funghi che nel cuocere induriscono, si facciano cuocere a lungo, mettendo nel tegame anche del salnitro. Si cuociano preferibilmente insieme alla carne (per diluirli) e con piccioli di pere selvatiche (diversamente questi ultimi potranno essere mangiati alla fine del pasto come antidoto).

Macrolepiota procera © Giuseppe Mazza
Si tenga presente che l’aceto è per sua natura contrario ai veleni, e pertanto va sempre usato senza parsimonia quando si mangiano i funghi ” (da notare come quest’ultimo consiglio, se seguito nella preparazione di Coprinus atramentarius, può provocare una intossicazione seria quale la sindrome coprinica ndr).
DAL MEDIOEVO AL SETTECENTO
Con la caduta dell’impero romano d’occidente e il conseguente imbarbarimento delle popolazioni, dovuto alle invasioni dei popoli nordici, si entra in quello che è chiamato “medioevo”. La cultura classica e scientifica passa nelle mani del clero e soprattutto dei monaci Benedettini che conservano le opere, le trascrivono e le traducono. Il medioevo è avaro di notizie e pochi sono gli studiosi di funghi che ci hanno tramandato le loro opere. È invece con il Rinascimento che si apre una nuova era per la botanica, che viene riconosciuta come vera scienza, anche se solo di complemento alla medicina. In questo periodo vengono rispolverati Dioscoride e Plinio con traduzioni, commentari e riedizioni.
ERMOLAO BARBARO (1454-1492) amante della cultura di Aristotele, Plinio e Dioscoride, tradusse e commentò le loro opere.
PIER ANDREA MATTIOLI (1500-1577) nato a Siena ma trentino di adozione, è stato un medico di grande fama. La sua opera principale è Commentari in Pedacii Dioscoridis Anarzabei de Materia medica, in essa viene riportata la traduzione dal greco al latino dell’opera originale di Dioscoride. A ogni capitolo è inserito, come appendice, un lungo commento molto più esteso del capitolo stesso. Fu un’opera di grande pregio tanto che in due secoli ebbe 60 edizioni, in originale, in italiano, in francese, in tedesco e in ceco. In quell’epoca, quando la comunità scientifica iniziò a considerare la botanica come una vera disciplina, anche se legata alla medicina, i funghi venivano comunque trattati marginalmente.

Cordoni miceliari nel sottosuolo © Giuseppe Mazza
Tuttavia il 1500 vide la nascita dei primi trattati monografici. Il primo di questi fu pubblicato da un medico olandese: JONGHE HADRIAN (1512-1575) che nel 1564 scrive una dissertazione su una nuova specie che chiamò Phallus impudicus, dal titolo “Phalli ex fungorum genere in Hollandiae sabuletis passim crescentis, descripto et ad vivum expressa pictura”, dove era descritto ed illustrato con una tavola xilografica, un fungo, che in seguito verrà chiamato in suo onore Phallus hadriani; che cresceva nei terreni sabbiosi in Olanda. L’opuscolo riportava anche le virtù medicinali del fungo considerato terapeutico per la gotta e dolori artritici.
PIER ANDREA CESALPINO (1525-1603) nato ad Arezzo nel 1525, quasi contemporaneo del Mattioli, anche lui medico celeberrimo, il Cesalpino ha lasciato una profonda impronta di sè nella botanica. In campo medico fu il primo a scoprire e descrivere in termini moderni la circolazione sanguigna e i capillari sanguigni, anche se il merito fu attribuito, ingiustamente, più tardi all’inglese Harvey.
Il Cesalpino fu medico personale di Papa Clemente VIII, fu archiatra pontificio e professore di medicina alla Sapienza di Roma, fu medico e amico di San Filippo Neri e Torquato Tasso. Il suo principale lavoro di botanica fu l’opera De Plantis in 16 libri, dove mette in atto un primo tentativo di catalogare le piante in maniera sistematica, secondo criteri precisi. Per questo è considerato il padre della botanica italiana. Il Cesalpino, da botanico, tratta ovviamente anche di funghi, è il primo che tenta di fare uno studio approfondito su di essi, ed è il primo che oltre alla descrizione delle varie specie, dà dei consigli su come cucinare quelli commestibili. Fin qui, la conoscenza dei funghi, si può dire, non aveva fatto grandi progressi. Pesava fortemente l’influenza delle antiche credenze, l’assenza di strumenti scientifici adatti lasciava vuoti incolmabili soprattutto sulle modalità della loro nascita, si credeva che i funghi fossero forme patologiche delle radici o degli alberi, se non addirittura che avessero origini magiche. Il progresso della botanica era stato notevole, molto meno quello dei funghi.
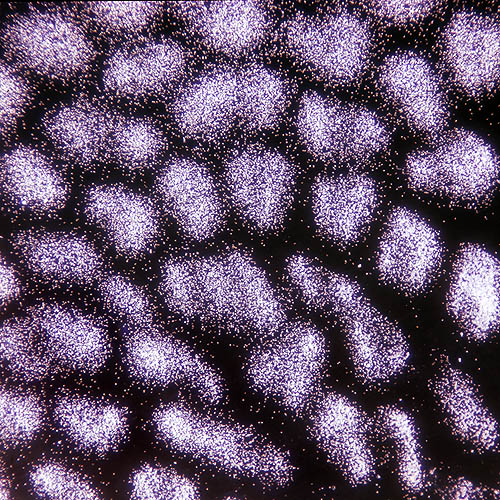
Spore di Boletus aestivalis al microscopio, a debole ingrandimento © Giuseppe Mazza
DAL SETTECENTO AI GIORNI NOSTRI
Con il settecento vi sono stati diversi scienziati che hanno parlato di funghi e hanno tentato di dare loro una sistemazione nel mondo della botanica, ma è con l’arrivo di PIETRO ANTONIO MICHELI (Firenze 1679-1737) che nasce la Micologia moderna, così come ancora oggi la conosciamo.
Nel Micheli la passione per la botanica nasce dalla lettura del Mattioli, del Bocconi e di altri scienziati, mentre lavorava come ragazzo di bottega presso un rilegatore di libri. Diventa un botanico di fama internazionale, pur senza avere alcun titolo di studio, spinto solo da una grande passione, aiutato ed erudito anche dai monaci vallombrosani, in particolare, per quanto riguarda la micologia, soprattutto da Padre Bruno Tozzi, grande botanico dell’epoca, socio della Royal Society di Londra.
Il Micheli è considerato, con giusto merito, il padre della micologia moderna. A lui va attribuito il grande merito di aver scoperto e dimostrato che anche i funghi si riproducono per seme, con la scoperta delle spore e la loro germinazione. Ferma era la sua convinzione che nessuna pianta si possa riprodurre senza seme, e che non esiste nessuna pianta priva di fiore e seme. Con l’ausilio di un rudimentale microscopio a tre lenti, dove per aumentare gli ingrandimenti era necessario cambiare fisicamente l’obiettivo, il Micheli riuscì a scoprire e descrivere i cheilocistidi ed i pleurocistidi. Così descrive quello che vede:
“ … dei minutissimi semolini distribuiti infra di loro, per quanto osservare potei, con ordine regolarissimo; e quello che mi fece più meravigliare fu il vedere che ognuno di loro stava situato sopra una base, la quale mi fece dubitando dire: chi sa che non sia il fiore o il calice dei funghi?”
Ancora:
“ … nell’orlo della lamella … nascono fiori apetali, nudi, costituiti da un semplice filamento cilindrico; in alcune specie questi fiori sono solitari, ossia disuniti, mentre in altre specie sono riuniti in masse o fiocchi. Sulla superficie della lamella nascono ovunque semi rotondi o tondeggianti, minutissimi, in alcune specie dispersi singolarmente, in altre specie riunite quattro per quattro”.
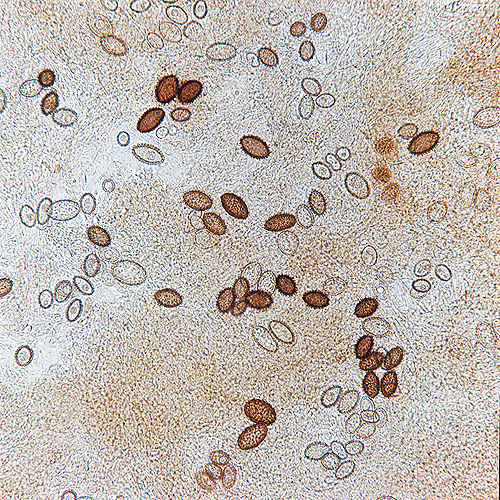
Spore di Tuber melanosporum osservate al microscopio © Giuseppe Mazza
“ … in alcune specie di funghi, e particolarmente in quelli che nascono sullo sterco equino, bovino e simili, abbiamo visto qualcosa degno do considerazione: e cioè che sulle facce delle lamelle si trovano non soltanto i semi, ma anche certi corpi diafani, di forma talora conica, talora piramidale, e che sembra essere stati qui collocati provvidamente dalla natura a impedire che le lamelle si tocchino con quelle adiacenti e ostacolino, trattenendoli fra di esse, la caduta dei semi giunti a maturazione …”.
La storia di questa affascinante materia si arricchisce notevolmente nell’800 e 900, anche grazie a nuove metodologie e nuovi strumenti, ma noi ci fermiamo qui, perché sarebbe troppo lungo raccontare tutta la storia della micologia dopo il Micheli. Infatti, dal settecento in poi abbiamo avuto grandi botanici, sistematici e micologi, da LINNEO a FRIES a PERSOON e tanti altri ancora, fino ad arrivare a DON GIACOMO BRESADOLA, il più grande micologo della fine 800 e inizio 900 che, senza mai spostarsi da Trento, ha studiato i funghi di tutto il mondo mediante le relazioni intrattenute con i più grandi scienziati del globo.
Tuttavia qualche riga in più va spesa per il grande Carl von Linné, nato il 23 maggio 1707 a Rashult, nella provincia di Smaland, in Svezia e morto a Uppsala nel gennaio del 1778, più noto come Linneo.
Nella sua opera più importante “ Systema naturae ” (1735) illustra per la prima volta il suo metodo di classificazione tassonomica dei regni animale, vegetale e minerale. Nel 1753 con l’opera “ Species Plantarum ” introduce la nomenclatura binomia dove ogni animale, ogni pianta e ogni minerale sono indicati con due nomi latini: il primo indica il genere mentre il secondo la specie.
La rivoluzione derivata dal sistema di Linneo fa si che nel 1867, a Parigi, venga organizzato il primo Congresso Internazionale di Botanica dove furono adottate le “Leggi di nomenclatura botanica”. È durante il secondo Congresso Internazionale di Botanica, tenutosi a Vienna nel 1905, che si adottano le “Regole Internazionali di Nomenclatura Botanica” (il primo codice). Da allora circa ogni 5 anni si indicono Congressi a livello mondiale per apportare eventuali modifiche, l’ultimo si è tenuto a Vienna nel 2005 e nel 2006 è stato pubblicato l’ultimo Codice Internazionale di Nomenclatura Botanica denominato (Vienna Code).
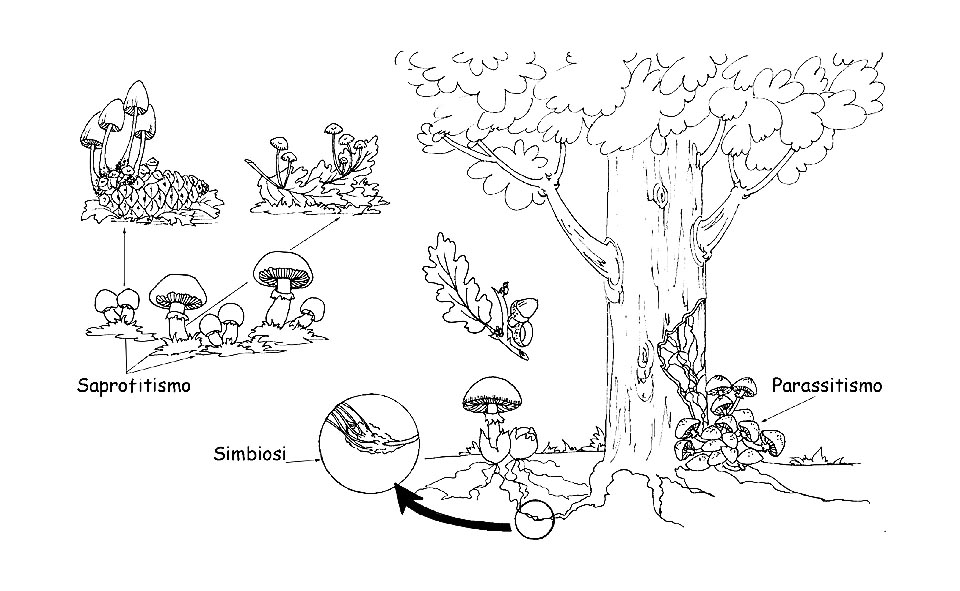
Modi di nutrizione dei funghi © Maria Tullii
IL FUNGO
Il fungo è un organismo vivente formato da cellule unite e comunicanti tra di loro, in una struttura costante, diversificate tra loro in funzione del loro fine biologico. Il vero organismo fungo è costituito da un insieme di filamenti (detti ife) che si diramano all’interno del substrato, spesso per parecchi metri. L’insieme di queste ife si chiama “micelio”. Singolarmente le ife non sono visibili ad occhio nudo ma si intrecciano tra loro fino a formare agglomerati miceliari: “cordoni miceliari” o “sclerozi”. Le ife si allungano nel terreno mediante la suddivisione della cellula apicale o di cellule intermedie, che danno origine a ramificazioni laterali. In questo modo il micelio aumenta la propria superficie e la propria capacità di assorbimento delle sostanze nutritive. Il fungo comunemente detto, quello che viene raccolto, altro non è che il frutto prodotto dal micelio quando le condizioni di umidità e temperatura sono favorevoli. Il frutto è più propriamente chiamato “carpoforo” o “sporoforo”. Anch’esso è costituito da agglomerati di ife. Come per tutti gli altri frutti la sua funzione è quella della riproduzione, che vedremo in dettaglio più avanti.
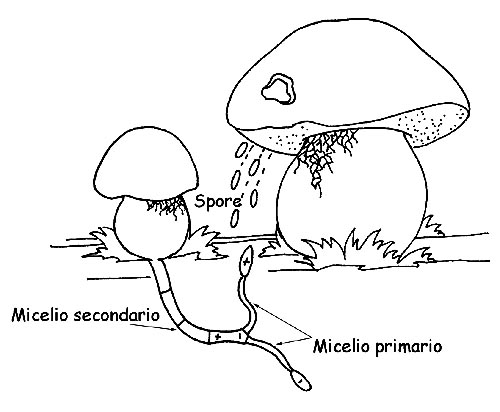
Ciclo riproduttivo dei funghi © Maria Tullii
L’HABITAT
I funghi nascono e crescono praticamente in qualsiasi habitat. Possono prediligere le zone aperte, o gli ambienti più umidi, un terreno acido o basico, la convivenza con essenze arboree o arbustive o praticole; insomma quale che sia la conformazione dell’habitat ci saranno una o più specie fungine adatte a colonizzarlo. Esistono inoltre specie che chiameremo “ubiquitarie”, in grado di colonizzare habitat diversi. L’habitat, dal canto suo, influisce sulla crescita e sull’aspetto dei carpofori: la stessa specie può presentare dimensioni e caratteri morfologici diversi a seconda dell’altitudine, della maggiore o minore umidità, della maggiore o minore esposizione ai raggi solari.
CONDIZIONI DI CRESCITA
Le condizioni di crescita dell’organismo fungo sono fortemente dipendenti dalle condizioni ambientali: quando umidità e temperatura sono favorevoli il micelio produce i carpofori, che si sviluppano e producono le spore che, a loro volta, cadendo nel terreno, daranno inizio ad un nuovo ciclo vitale. Le condizioni di crescita variano da zona a zona: il periodo di crescita fungina sarà più breve laddove il clima è più rigido e la stagione invernale è prolungata, quasi nullo nelle regioni a carattere permanente di siccità, pressoché ininterrotto nelle regioni a clima temperato.
LA RIPRODUZIONE
Come si diceva, il micelio, in condizioni climatiche di umidità e temperatura opportune, fruttifica, dando origine ai carpofori. Per riprodursi, mediante cellule specializzate, producono milioni di semi detti “spore” che, disperse nell’ambiente circostante con l’ausilio di fattori esterni quali vento, pioggia o vettori animali, anche a grande distanza, danno origine ad un nuovo ciclo vitale. Le spore sono cellule piccolissime, non visibili ad occhio nudo, osservabili quindi solo al microscopio. L’unità di misura delle spore è il micron (µm), che corrisponde a un millesimo di millimetro. Le spore sono sessuate (almeno nei funghi superiori, quelli di cui stiamo trattando), non nel senso comune del termine (maschile o femminile) ma per la diversa carica che possiedono: positiva (+) o negativa (-).
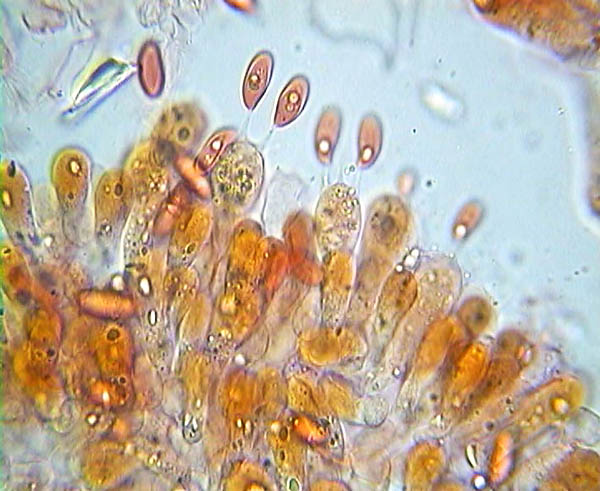
Basidi e spore di Lepiota alba © Pierluigi Angeli
Quando una spora trova un substrato adatto alla riproduzione e in condizioni favorevoli, germina, dando origine ad un filamento con la stessa carica della spora originaria (ifa), che può svilupparsi e crescere (micelio primario), ma non può fruttificare.
Perché il micelio primario possa fruttificare, deve combinarsi con un’altro micelio primario della stessa specie ma di segno opposto.
L’unione dei due miceli, negativo e positivo, è detto micelio secondario, l’organismo “fungo” vero e proprio. Tuttavia, la possibilità che dal micelio primario si formi una nuova colonia fungina è piuttosto rara ed è per questo che ciascun corpo fruttifero rilascia nell’ ambiente milioni e milioni di spore.
SISTEMI DI NUTRIZIONE
Tutte le forme di vita presenti sulla Terra si dividono in :
PRODUTTORI o AUTOTROFI : organismi viventi in grado di nutrirsi in modo autonomo sintetizzando le sostanze di cui hanno bisogno partendo dai composti inorganici presenti in natura: le piante usano la luce solare per produrre dei composti complessi (come gli zuccheri) a partire da composti semplici come anidride carbonica ed acqua, attraverso il processo della Fotosintesi clorofilliana. CONSUMATORI o ETEROTROFI : organismi viventi che si procurano le sostanze necessarie per la loro sopravvivenza dai produttori o da altri consumatori. Le fonti da cui viene ottenuto il cibo, permettono di suddividere i consumatori in quattro gruppi: Erbivori: si nutrono di sostanze vegetali; Carnivori: si nutrono di altri consumatori; Onnivori: si nutrono tanto di sostanze vegetali che di altri consumatori; Decompositori: si nutrono utilizzando gli organismi morti o viventi, trasformando le sostanze organiche complesse in sostanze semplici, come anidride carbonica ed acqua. Le piante superiori, essendo in grado di autoalimentarsi, trasformando le sostanze inorganiche (acqua e anidride carbonica) in composti complessi, mediante il processo della fotosintesi clorofilliana, appartengono al gruppo degli organismi Autotrofi. I funghi, un tempo classificati tra i vegetali, appartengono ora ad un Regno proprio: il Regno Fungi, in quanto non sono in grado di svolgere la funzione clorofilliana. Sono perciò da annoverarsi tra gli organismi Eterotrofi e tra questi ai Decompositori.
In base al sistema nutrizionale adottato, i funghi si classificano quindi in :

Clitocybe odora, un fungo decompositore della lettiera © Giuseppe Mazza
– Saprofiti: si nutrono di sostanze organiche in decomposizione, sia vegetali che animali. Realizzano una prima fase di disgregazione e preparano il terreno ai batteri che completeranno il ciclo, così che tutti i residui organici vengano ritrasformati in sostanze inorganiche: acqua, anidride carbonica e sali minerali. Fanno parte di questo gruppo i funghi che nascono su prati e pascoli, foglie e rami secchi o organismi animali morti. Possiamo considerare i funghi saprofiti come operatori ecologici della natura: senza di loro i detriti vegetali e animali soffocherebbero l’ambiente.
– Parassiti: vivono a spese di altri esseri viventi, sia vegetali che animali. Se, da un lato, i funghi parassiti rappresentano un vero pericolo per piante e animali, perché provocano serie patologie fino a causarne la morte, dall’altro fungono da elemento regolatore, sopprimendo le piante più deboli o malate, favorendo lo sviluppo di quelle sane. Una volta morto l’organismo ospite, vegetale o animale che fosse, il fungo parassita si trasforma in saprofita, finendo di demolire la sostanza organica morta.
– Micorrizici o simbionti : sono i funghi che vivono in simbiosi con le piante superiori, stabilendo un rapporto di mutuo vantaggio. Le parti apicali del micelio avvolgono la parte terminale dei peli radicali delle piante superiori, aumentandone la superficie d’assorbimento. La pianta ha così a disposizione una maggior quantità di sostanze inorganiche, che trasformerà, mediante la fotosintesi clorofilliana, in composti organici che mette a disposizione del fungo. Inoltre la “micorriza” (formata dai peli radicali avvolti dalle ife fungine) protegge le radici della pianta dagli attacchi patogeni, rendendola più forte e migliorandone la salute. Le micorrize possono essere: Endomicorrize, quando le ife fungine penetrano negli spazi intercellulari e all’interno delle cellule vegetali. Il fungo si espande attraverso le cellule corticali, tuttavia senza intaccare il cilindro centrale e le cellule apicali della radice. Ectomicorrize, quando le ife fungine circondano la radice penetrando negli spazi intercellulari ma non all’interno delle cellule vegetali. L’intreccio delle ife attorno alle cellule radicali è detto reticolo di Hartig.
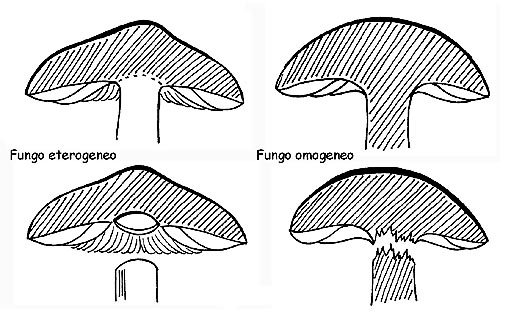
Omogeneità del caproforo © Maria Tullii
MORFOLOGIA DEI FUNGHI
Perché ad un fungo possa correttamente essere assegnato un cognome ed un nome (Genere, specie) occorre osservarne tutti i caratteri.
Cominciamo a distinguere alcune tipologie fungine:
Agaricomiceti : sono tutti quei funghi composti da cappello e gambo.
– Ascomiceti: si presentano a volte con pseudogambo e pseudocappello, oppure a forma di coppa, sessili o non, oppure in forma di tubero e in questo caso, ipogei o semiipogei.
– Gasteromiceti: in forma ± rotonda, con o senza pseudogambo, ipogei o epigei.
Prendiamo ora in esame i caratteri morfologici che vanno osservati nei funghi composti da cappello e gambo: le differenze, talvolta, sono minime e solo un’osservazione accurata ne consente l’esatta determinazione.
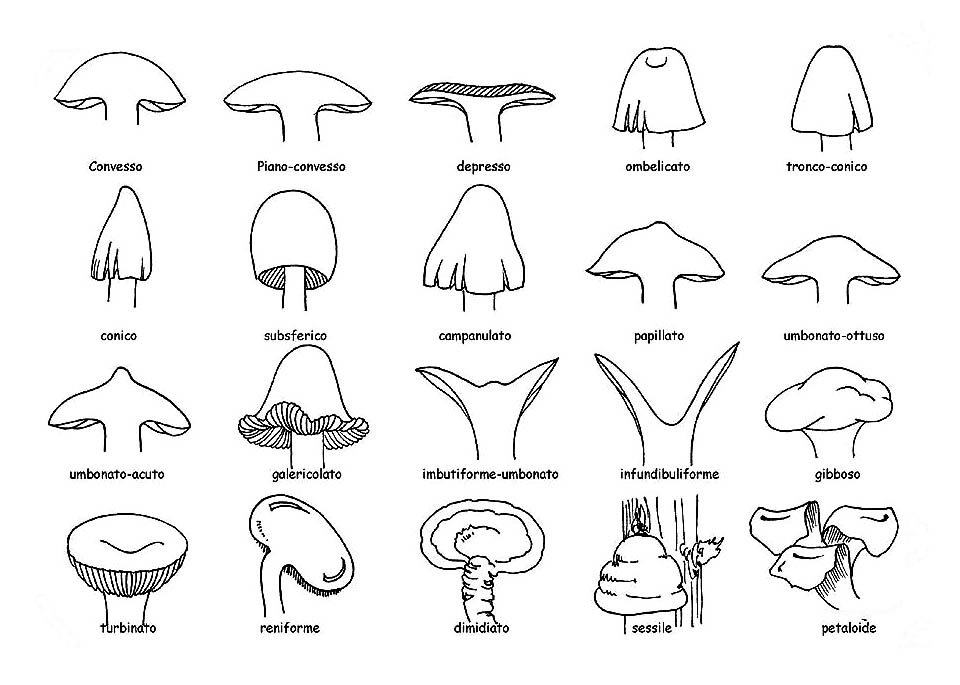
Forma del cappello © Maria Tullii

Margine e profilo del cappello © Maria Tullii
OMOGENEITA’
Il carpoforo è costituito da un intreccio di ife strettamente unite e disposte per lo più parallelamente fra di loro. Se queste ife hanno la stessa struttura sia nel cappello che nel gambo, avremo un fungo omogeneo. Questa caratteristica è facilmente verificabile: se quando si tenta di separare il cappello dal gambo, si procura la rottura della carne. Al contrario, se le ife sono strutturate in modo diverso nel cappello e nel gambo,avremo un fungo eterogeneo e pertanto la separazione di queste due parti avverrà senza rottura, come se le due parti fossero semplicemente appoggiate l’una sull’altra.
CAPPELLO
Del cappello si osservano:
– le dimensioni, corrispondenti al suo diametro misurato a completo sviluppo.
– la forma che varia notevolmente dallo stadio giovanile a quello maturo. Normalmente, si prendono in considerazione gli esemplari adulti, ma sarebbe meglio osservare più esemplari, a diversi stadi di crescita. Negli esemplari giovani, di solito, il cappello è più chiuso per poi aprirsi ed appianarsi fino a raggiungere, nell’adulto, la forma tipica.
– il margine, sempre importante, addirittura determinante in alcune specie, va attentamente osservato con riferimento al piano orizzontale del cappello. Può presentarsi diritto e quindi sullo stesso piano; involuto, arrotolato in dentro; revoluto, arrotolato in fuori; ricurvo, piegato verso il basso; il profilo del margine può presentarsi: regolare, ondulato, lobato, festonato, sinuoso o plissettato, intero o striato; è inoltre considerato eccedente quando sporge, anche di poco, oltre l’imenio (lamelle, tubuli, ecc.).

Decorazioni del cappello © Maria Tullii
la cuticola del cappello, può avere diversi aspetti: liscia, viscida o viscosa, tomentosa, pelosa, vellutata o ruvida. Osserviamo, inoltre, se può essere separata dalla carne del cappello: la cuticola che non viene via si dice adnata o non separabile, se, al contrario, viene via interamente si dice che è separabile, se si separa solo in parte, diremo che è parzialmente separabile e osserveremo la porzione di raggio del cappello che viene scoperta (1/2, 1/3, ecc.).
– la superficie del cappello, può o non presentare decorazioni. Una superficie priva di decorazioni può essere unita, glabra, uniforme; ove presenti le decorazioni possono essere:
Innate, ossia parte integrante della struttura della pellicola, come fibrille, vergature o macchie.
Adnate o applicate, derivanti cioè da dissociazioni della pellicola come screpolature e squame, oppure residui del velo generale quali verruche, placche, farinosità, villosità, pruinosità.
– il colore è un carattere importante ma non assoluto, in quanto la maggior parte dei funghi presenta una variabilità cromatica notevole in rapporto alle condizioni climatiche, all’età, all’esposizione. Per descrivere un colore faremo riferimento a colori comunemente conosciuti e associati ad oggetti quotidiani (rosso mattone, verde oliva etc).
IMENOFORO
L’imenoforo è l’insieme delle strutture macroscopiche e microscopiche preposte alla produzione delle spore.
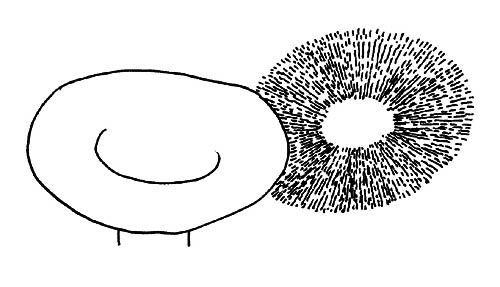
Sporata di massa © Maria Tullii
Esso riveste importanza fondamentale, non solo dal punto di vista biologico, ma anche da quello della determinazione.
Nei funghi a gambo e cappello è generalmente situato nella zona inferiore del cappello e può essere formato da lamelle, tubuli e pori, aculei; in alcuni generi è quasi liscio o appena rugoso.
Esposto all’aria nella maggior parte dei casi, l’imenio è invece protetto e racchiuso all’interno del carpoforo nei funghi appartenenti alle Gastromycetidae (le spore verranno liberate, a maturazione del carpoforo, per rottura del peridio) e, tra gli ascomiceti, alle Tuberales.
Le lamelle il primo carattere che dobbiamo osservare è il colore delle lamelle che può essere: innato, cioè il colore visibile è proprio della lamella; derivato, cioè il colore della lamella deriva dalla colorazione delle spore.
Carattere questo di enorme importanza in quanto l’appartenenza ad un gruppo cromosporeo determina l’assegnazione di un fungo ad un gruppo di generi o ad un altro. I gruppi cromosporei sono cinque: leucosporei, cioè a sporata bianca o biancastra; ocrosporei, cioè a sporata ocra; rhodosporei, cioè a sporata rosa; iantinosporei, cioè a sporata viola; melanosporei, cioè a sporata nera.
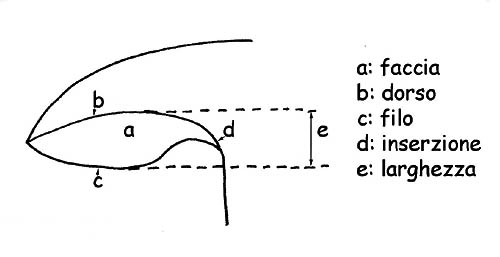
Parti della lamella © Maria Tullii
Il colore della sporata è verificabile tramite la raccolta della polvere sporale su supporto di colore contrastante. Osserveremo poi l’omogeneità delle lamelle.
Analogamente a quanto già detto per il cappello e per la cuticola osserveremo se le lamelle si staccano facilmente dalla carne del cappello: se si diremo che sono eterogenee o separabili; se non si staccano ma si rompono nel tentativo, diremo che sono omogenee o non separabili.
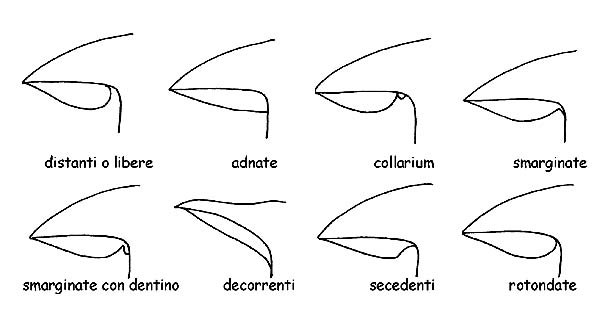
La forma delle lamelle in tutte le sue componenti: orlo, filo, larghezza, spessore, ecc., l’ inserzione sul gambo, per cui vengono definite:
– libere o distanti, quando le lamelle non arrivano a toccare il gambo, caratteristica comune a tutti i funghi eterogenei.
– adnate, quando le lamelle si attaccano al gambo su tutta la loro larghezza.
Insezione delle lamelle © Maria Tullii
– smarginate o uncinate, quando si attaccano al gambo solo con una piccola parte della loro larghezza.
– decorrenti, quando proseguono lungo il gambo, oltre la loro larghezza.
– decorrenti per un dentino, quando la parte che decorre forma un piccolo prolungamento (dentino).
– collarium, quando le lamelle, distanti dal gambo, sono collegate ad esso mediante un piccolo cordone circolare.
Osservando il profilo lamellare all’orlo, potremo quindi definirle acute, sinuose, ventricose, ottuse o diritte.
Il dorso è lo spigolo lamellare attaccato alla carne del cappello.

Profolo lamelle © Maria Tullii
Il filo lamellare (che è lo spigolo opposto al dorso) può essere intero, seghettato, denticolato, liscio, fioccoso o pruinoso e può essere poi concolore alle lamelle o di colore diverso.
In questo caso parleremo di filo eteromorfo.
Se invece il filo lamellare si divide in prossimità del gambo, le lamelle si dicono forcate.
E bisognerà constatare la presenza o meno di Lamellule (sono così definite quelle lamelle che partono dal margine del cappello e non arrivano al gambo) che sono normalmente intercalate alle lamelle e hanno lunghezza variabile.
La parte terminale delle lamellule può essere tronca, rotondata o sinuata.
E occorre controllare anche la presenza o meno di Anastomosi: si tratta di congiunzioni trasversali che uniscono in più punti una lamella all’altra.
Per maggior chiarezza: per larghezza si intende la distanza tra il dorso ed il filo lamellare.
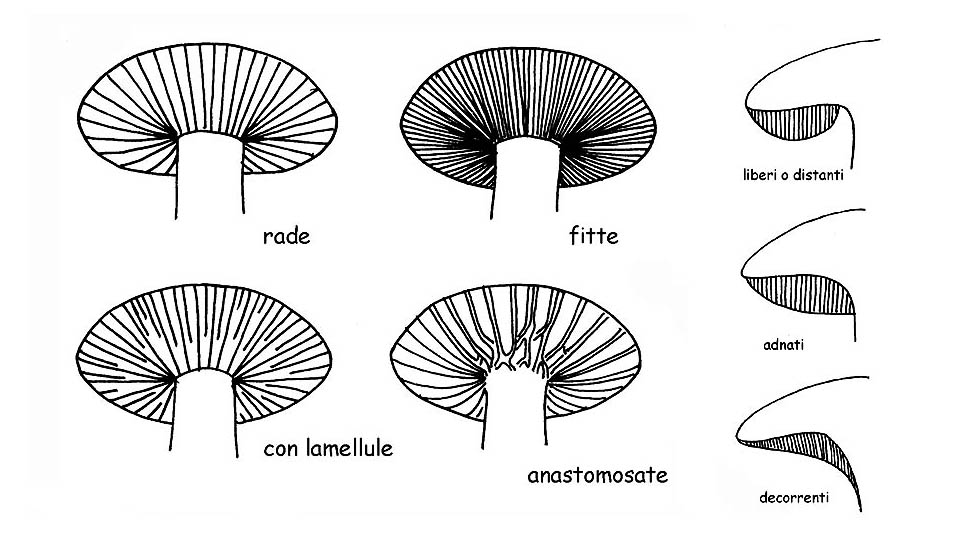
Tipologia lamelle e tuboli © Maria Tullii
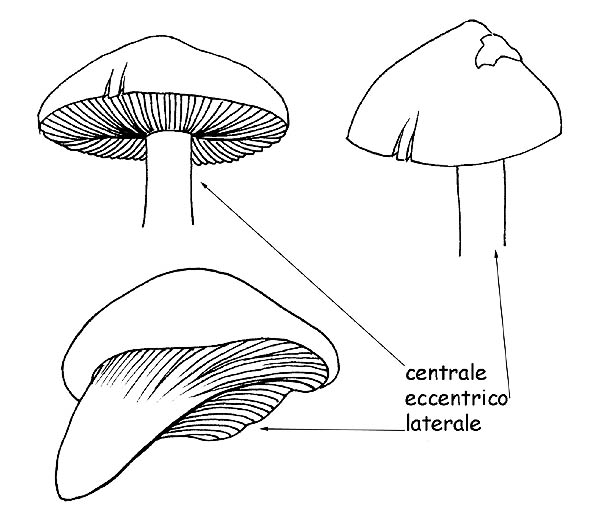
Inserzione del gambo © Maria Tullii
Con la definizione spessa o sottile ci si riferisce allo spessore della lamella.
Con le definizioni rade, fitte ci si riferisce alla distanza tra una lamella e l’altra.
Analogamente ai caratteri indicati per le lamelle, coi debiti aggiustamenti, osserveremo i caratteri dell’imenio a Tubuli e i pori.
Osserveremo il colore dei tubuli e dei pori nel fungo giovane e nel fungo maturo (tubuli e pori possono essere concolori ma anche di colore diverso), con tutte le variazioni intermedie; la lunghezza di tubuli, la loro attaccatura al gambo, la grandezza dei pori, la loro forma etc..
Nota bene: negli imenofori a tubuli e pori l’ omogeneità è un carattere che determina l’appartenenza ad un determinato gruppo: nelle Boletales la parte imeniale è eterogenea, quindi facilmente separabile; nelle Poliporales la parte imeniale è saldamente attaccata alla carne, quindi non separabile.
GAMBO
Se il carpoforo si presenta a cappello e gambo è definito peduncolato; quando il gambo non c’è è definito sessile.
Anche per il gambo i caratteri da osservare sono molti e tutti rivestono importanza fondamentale per la determinazione della specie.
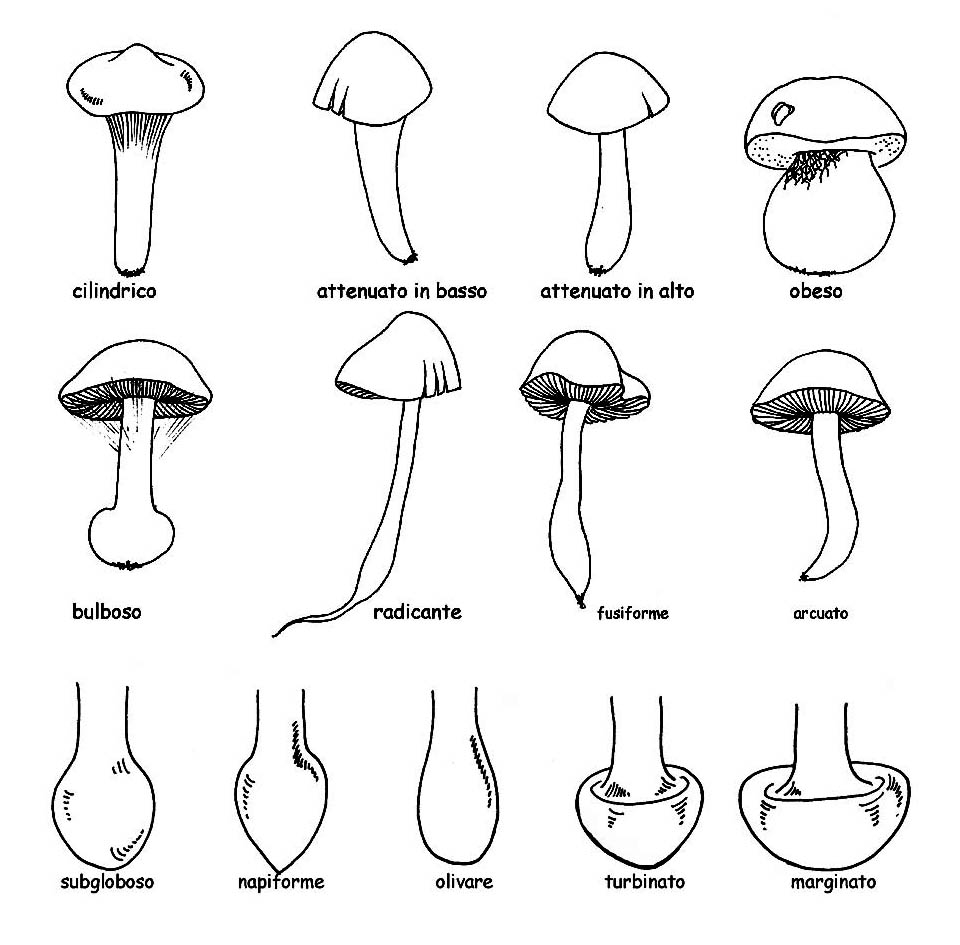
Forma del gambo e tipologia del piede © Maria Tullii
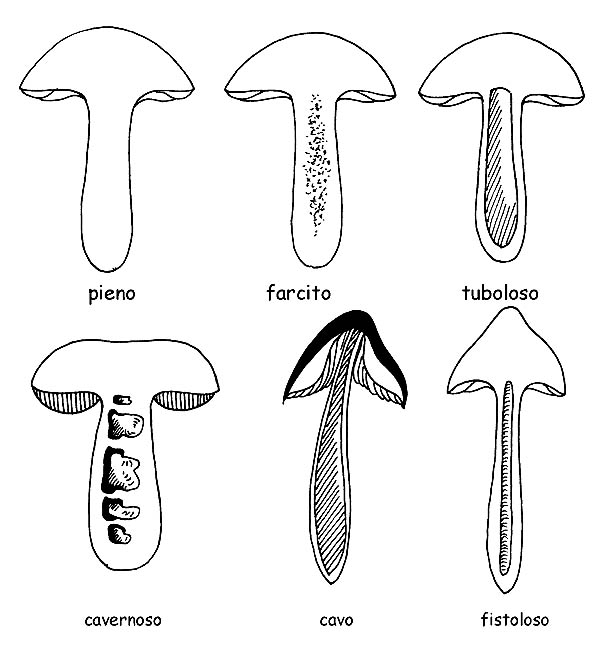
Interno del gambo © Maria Tullii
Ossereveremo:
– l’inserzione al cappello può essere centrale, laterale o eccentrica;
– la forma: cilindrica, attenuata in basso o in alto, obesa, ventricosa o bulbosa.
– il bulbo alla base può essere definito, secondo la forma, subsferico, napiforme, olivare, turbinato e marginato.
– l’interno può presentarsi carnoso, fibroso-carnoso, fibroso, pieno o cavo, fistoloso, corticato etc.
– Possono essere presenti decorazioni di vario tipo, per cui la superficie può presentarsi liscia, fioccosa, squamosa, fibrillosa, verrucosa, pelosa, punteggiata e calzata, o diversamente decorata; ma anche secca, vischiosa o glutinosa.
Un discorso a parte va fatto per il gruppo delle Boletaceae ove le decorazioni del gambo sono rappresentate per lo più da un reticolo. In questo caso vanno osservati: il colore, la grandezza delle maglie, se queste sono ± allungate o se sono ± in rilievo.
Il reticolo può essere anche totalmente assente e il gambo può infine presentarsi ricoperto da punteggiature, fibrille, asperità o scaglie come nel genere Leccinum.
VELI
Tutti i carpofori, nello stadio iniziale della vita (primordio) sono avvolti in una membrana chiusa che li avvolge (velo generale).
Nella maggior parte dei funghi questo velo scompare già nei primi stadi di sviluppo, ma in altri generi, appartenenti alle Agaricaceae , il velo generale persiste fino all’età adulta ed è riscontrabile sul carpoforo sotto varie forme: verruche o placche sul cappello, volva o armilla alla base del gambo; sempre nelle Agaricaceae , alcuni generi presentano un altro velo: il velo parziale, che copre la parte inferione del cappello, partendo dal margine del cappello e arrivando fino al gambo, a protezione dell’imenoforo.
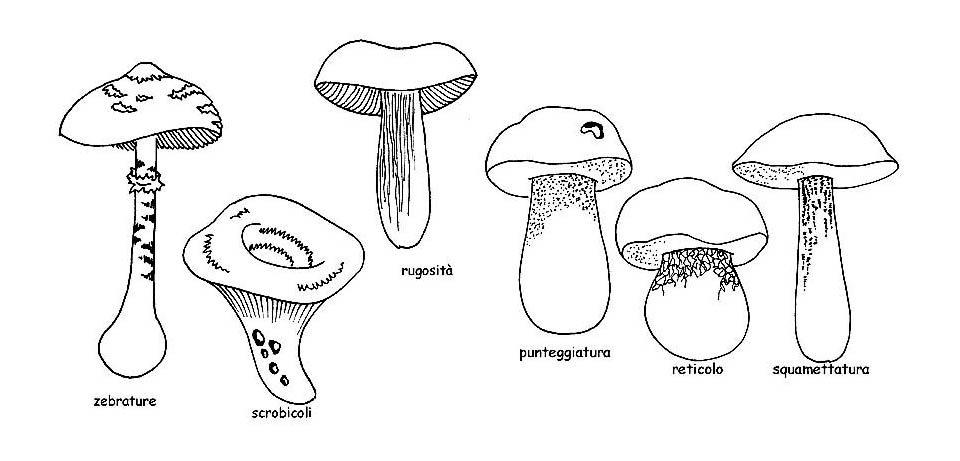
Decorazioni del gambo © Maria Tullii
Questo velo dapprima chiuso negli esemplari giovani, si stacca dal margine del cappello a mano a mano che il cappello si apre, restando attaccato al gambo dove rimane, ± evidente, sotto forma di anello.
La volva può essere membranosa, a sacco, inguainante, circoncisa, dissociata in anelli o in perle; l’anello può essere fioccoso, largo, stretto, infero (quando si stacca dall’alto verso il basso), supero (quando si stacca dal basso verso l’alto) ecc.. se residuo del velo generale si chiamerà armilla. In alcune specie è presente un doppio anello, residuo del velo generale e del velo parziale uniti tra loro sul gambo. In altre il residuo del velo parziale è rappresentato da filamenti sottilissimi e fugaci (cortina).
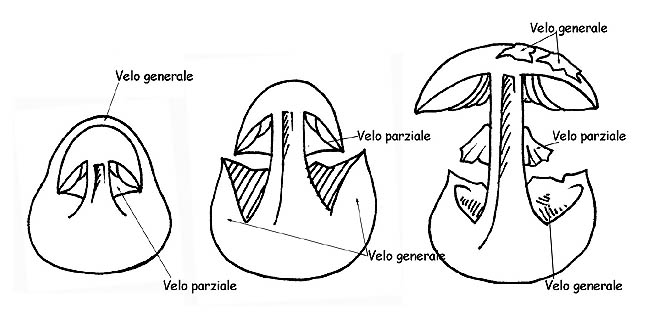
Sviluppo dei veli © Maria Tullii
CARNE
La polpa di un fungo, cioè la parte che si osserva tagliando il fungo dall’alto verso il basso si chiama carne. Analogamente alle altre parti del fungo, anche la carne presenta dei caratteri fondamentali per la determinazione della specie. A questo proposito la consistenza della carne può essere soda, compatta, molle, coriacea, membranosa, ceracea, ecc. Va poi considerato il colore ed eventualmente il viraggio, cioè la mutazione del colore della carne esposta all’aria, dovuta all’ossidazione di due enzimi, carattere questo (particolarmente importante nelle Boletaceae) che va osservato con attenzione, annotando l’ intensità ed il tempo di comparsa del viraggio. Può avere tonalità blu o rosse, secondo la dominanza di un enzima sull’altro. L’intensità del viraggio potrà essere maggiore negli esemplari giovani e freschi o più debole o inesistente negli esemplari più vecchi; l’ igrofaneità: è la capacità ± evidente della carne di assorbire acqua; la reviviscenza: è la capacità della carne di essiccare e tornare fresca al contatto con l’acqua; i caratteri organolettici: odore e sapore della carne sono caratteri importanti, addirittura determinanti in alcune specie del genere Agaricus.
CENNI DI SISTEMATICA
La micologia è una ramo delle Scienze Naturali, quello che tratta dei funghi. Come ogni scienza, per essere chiara, deve avvalersi di strumenti di ricerca e catalogazione. Eccoli:
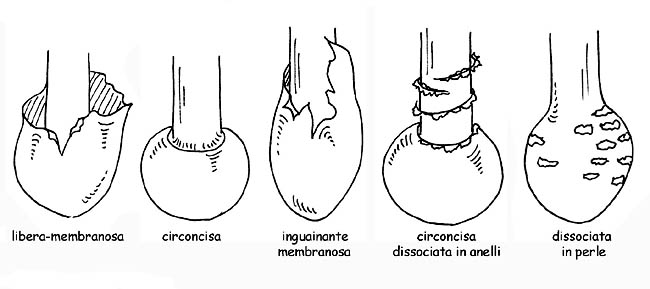
Tipologie di volva © Maria Tullii
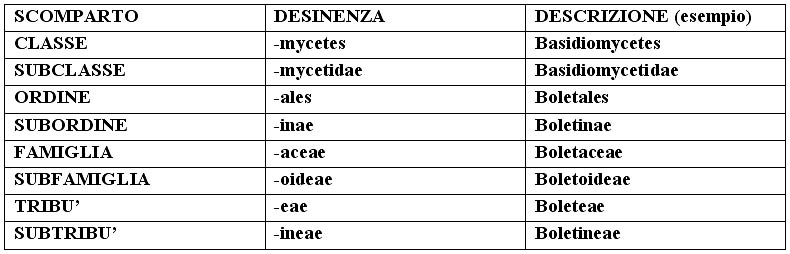
– la SISTEMATICA: è lo strumento scientifico che permette di sistemare l’oggetto di studio entro uno schema preciso (sistema) caratterizzato da una suddivisione in scomparti, ciascuno con caratteristiche comuni, all’interno di ognuno dei quali siano presenti altri scomparti, via via a scendere fino ad arrivare all’unità (una sorta di scatole cinesi dove la specie è contenuta all’interno di un genere, il genere all’interno di una famiglia ecc. vedi schema in calce). È uno strumento in continuo aggiornamento e non univoco, in quanto può essere interpretato in modo diverso dai singoli micologi. La classificazione che proponiamo è quella che ci deriva dai padri della sistematica moderna.
– la TASSONOMIA è la scienza che studia la classificazione dell’oggetto di studio, ossia il corretto posizionamento dei vari taxa nel disegno sistematico. Per taxon si intende un raggruppamento, di qualsiasi grado (specie, genere, famiglia, ordine, classe, divisione, regno).
– la NOMENCLATURA, è l’insieme delle regole che consente l’assegnazione di un nome scientificamente valido e corretto all’oggetto di studio. Queste regole sono sancite dal Codice Internazionale di Nomenclatura Botanica (C.I.d.N.B.). A questo codice fanno riferimento tutti gli studiosi e gli autori di libri e pubblicazioni varie di botanica e micologia. Il C.I.d.N.B. è stato fondato nel 1867; ogni 5-6 anni si indicono Congressi e Comitati di livello mondiale per apportare aggiornamenti ed eventuali modifiche.
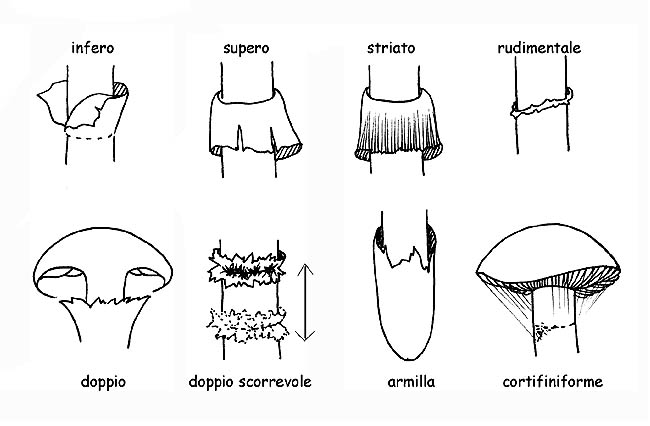
Tipologie di anello © Maria Tullii
CLASSIFICAZIONE E SCHEMA SISTEMATICO DEI FUNGHI
Proponiamo qui di seguito uno schema sistematico riguardante la classificazione dei funghi, riferendoci agli insegnamenti derivanti dalle scuole di R. Kühner e R. Singer, famosi studiosi di micologia e secondo i dettami del C.I.d.N.B (Codice Internazionale di Nomenclatura Botanica).
Nota: quando si cita una specie, il nome di questa va scritto nella forma binomia latina; il primo nome, quello del Genere, deve avere l’iniziale maiuscola (es. Agaricus), il secondo, quello della specie, minuscola (es. campestris); segue il nome, di solito abbreviato, dell’autore che ha determinato e descritto il fungo (es. Bull. per Bulliard); se (all’interno di una parentesi), dopo questo nome ce n’è un altro, può essere quello del sanzionatore, e in questo caso è sempre preceduto da “:”, e può essere solo Persoon o Fries, (es. Bull. : Fr.), oppure di chi ha completato la diagnosi originale, in caso di mancanze dell’autore (per esempio l’assenza della diagnosi latina), e in questo caso il nome è preceduto da “ex” (es. Bull. ex Vent.). Può poi essere presente un terzo nome (fuori della parentesi), che rappresenta l’autore che ha modificato la posizione del Genere o della specie (es. Bull.: Fr.) Kumm. La desinenza identifica i vari scomparti della sistematica. Il Genere, il sottogenere, la sezione, la specie, la varietà e la forma hanno desinenze variabili, legate alla radice linguistica. Fanno eccezione Regno e Divisione, che non hanno particolari desinenze. Dalla Classe alla Sottotribù, abbiamo in ordine discendente:
MICROSCOPIA
Come si è visto per i caratteri macroscopici, anche i caratteri microscopici dei funghi rivestono importanza fondamentale per la corretta identificazione della specie. In alcuni generi la microscopia è l’unico strumento certo per la determinazione di molte specie (ad es. nel Genere Mycena). In questa, che vuole essere una dissertazione divulgativa e non strettamente scientifica, illustreremo le principali strutture microscopiche presenti nei funghi superiori, i quali si dividono in due grandi classi: Ascomycetes e Basidiomycetes. Illustreremo in breve anche i reagenti e le tecniche più comunemente usati per la loro osservazione.
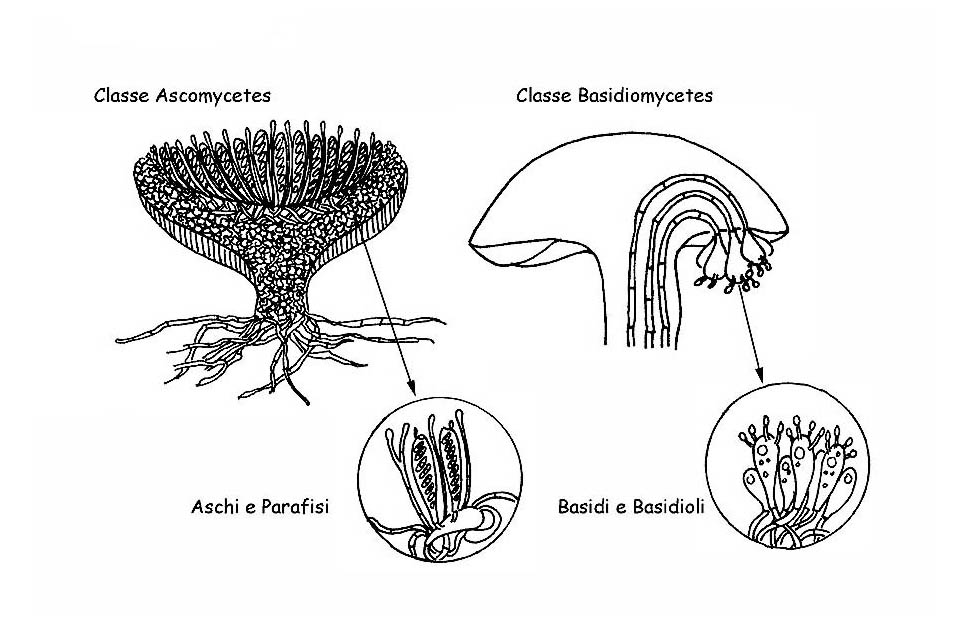
Funghi superiori © Maria Tullii
ASCOMICETI: I LORO ELEMENTI MICROSCOPICI
Ascomiceti © Maria Tullii
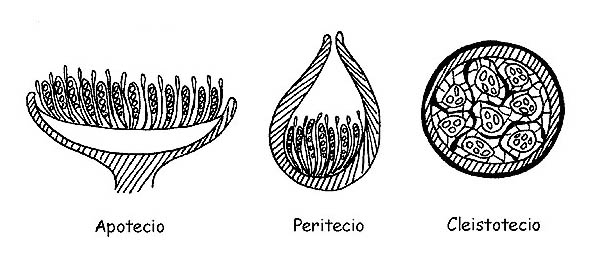
È importante, per questa classe di carpofori, premettere un cenno sullo sviluppo della parte fertile del fungo: l’imenio (tecio).
Quando il carpoforo espone completamente all’aria la sua parte imeniale diremo che siamo in presenza di un Apotecio, quando la parte imeniale comunica con l’esterno solo attraverso una piccola apertura apicale diremo che si tratta di un Peritecio e quando infine la parte imeniale non viene a contatto con l’esterno diremo che si tratta di un Cleistotecio (in questo caso la maggior parte delle specie hanno sviluppo ipogeo o semiipogeo).
Vedremo che questa distinzione è importante da un punto di vista microscopico per l’osservazione specifica dei vari elementi.
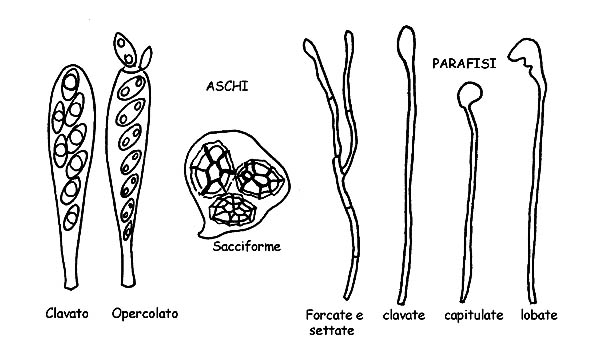
Aschi e Parafisi © Maria Tullii
Gli Aschi, da cui deriva la denominazione della classe, sono gli elementi fertili che producono le spore e si sviluppano da particolari ife specializzate, dette ascogene.
Si presentano cilindrici ± claviformi o in forma di otre o di sacco e contengono al loro interno le spore (per lo più in numero di otto, ma anche in numero variabile, fino a trentadue e oltre) fino a maturazione.
Possono avere parete singola o doppia (tunicati), possono avere reazione positiva o negativa al reattivo di Melzer, in caso di reazione positiva di dicono amiloidi o iodio positivi.
Gli aschi cilindroidi, tipici degli Apoteci e dei Periteci, si aprono, a maturazione delle spore, per lacerazione o spaccatura della parte apicale o mediante l’apertura di un minuscolo coperchietto (opercolo) lasciando fuoriuscire le spore mature.
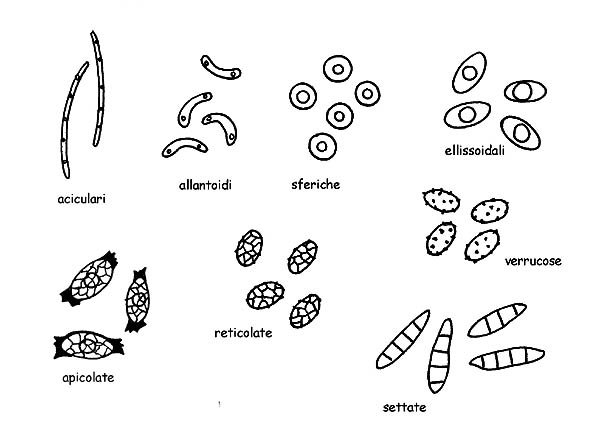
Spore © Maria Tullii
Gli aschi sacciformi tipici invece dei Cleistoteci, liberano le spore mediante la decomposizione delle pareti.
Le Parafisi sono sottili ife sterili, sono frammiste agli aschi e svolgono una funzione di sostegno per questi ultimi.
Si presentano per lo più cilindracee, possono essere forcate o settate, e l’apice può presentarsi in forme diverse: clavato, lanceolato, capitolato, lobato, incrostato ecc.
Anche nelle parafisi, per l’identificazione di alcune specie, si osserva la positività al reattivo di Melzer.
Le Spore negli ascomiceti assumono una grande varietà di forme (aciculare, allantoide, sferica, ellissoidale, apicolata ecc.), colori ed ornamentazioni (reticoli, creste, verruche, aculei ecc.).
Possono inoltre essere settate, sia longitudinalmente che trasversalmente.
I Peli sono elemento differenziale per molte specie di ascomiceti, proteggono l’imenoforo e trattengono l’umidità necessaria allo sviluppo della specie.
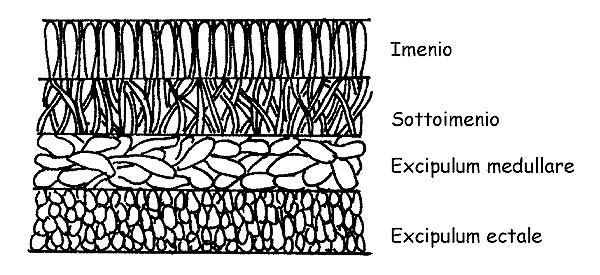
Sezione dell’Apotecio © Maria Tullii
Possono essere dislocati sulla superficie esterna o sul margine del carpoforo, presentano una grande variabilità di forma e di dimensioni, possono essere settati o non, con apice acuto o ± arrotondato ecc.
La Textura : per textura si intende la struttura della carne che può essere formata da cellule di diversa forma e disposizione.
È inoltre importante negli Apoteci, osservare la sezione completa del carpoforo.
Essa è costituita da più strati: Imenio, Sottoimenio, Excipulum medullare, Excipulum ectale.
L’osservazione della conformazione delle ife dei singoli strati può essere determinante per arrivare alla determinazione di una specie.
BASIDIOMICETI: I LORO ELEMENTI MICROSCOPICI
Negli ascomiceti abbiamo visto che le spore si sviluppano all’interno di strutture specializzate, gli aschi, che le proteggono fino a completa maturazione. Al contrario i basidiomiceti sviluppano e maturano le loro spore all’esterno di strutture fertili, i Basidi.
La classe Basidiomycetes si divide in due grandi sottoclassi: Holobasidiomycetes e Heterobasidiomycetes. Appartengono alla prima tutte le specie con basidi monocellulari e alla seconda le specie con basidi pluricellulari.
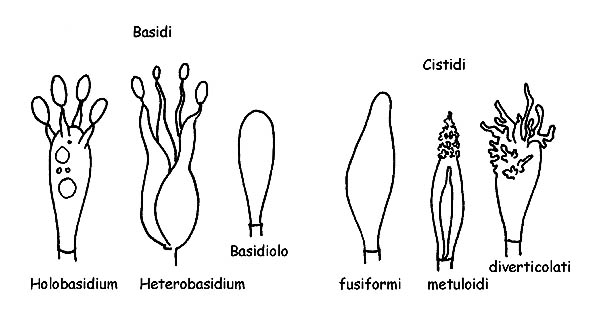
Strutture dei Basidiomiceti © Maria Tulliii
I Basidi sono cellule specializzate per la produzione delle spore. Nascono dalla cellula terminale di una ifa. All’interno del basidio avviene la divisione cellulare che da origine a quattro cellule ognuna delle quali diventerà una spora. La formazione delle spore avviene in cima a delle protuberanze poste all’apice del basidio (sterigmi) e, a maturazione, le spore verranno rilasciate nell’ambiente. I basidi sono generalmente ± cilindrici e ± clavati, presentano generalmente quattro sterigmi (basidi tetrasporici) ma possono anche presentarne due o tre o più di quattro, possono o no presentare del pigmento cellulare.
I Basidioli altro non sono che basidi ancora in formazione.
I Cistidi sono elementi sterili e possono essere presenti sia nella parte imeniale che sulla cuticola o sul gambo. I cistidi presenti sulle lamelle si chiamano: Pleurocistidi se presenti sulle facce lamellari, Cheilocistidi se presenti sul filo della lamella, mentre quelli della cuticola e del gambo si chiamano rispettivamente Dermatocistidi e Caulocistidi. I cistidi possono presentarsi in forme molto differenziate da specie a specie, con o senza incrostazioni, con parete semplice o doppia, con o senza pigmento intracellulare. Rivestono quindi grande importanza da un punto di vista microscopico in quanto consentono spesso una corretta determinazione.
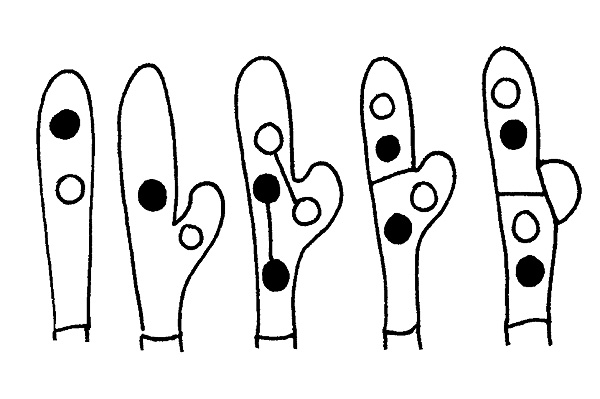
Formazione di un giunto a fibbia © Maria Tullii
Le Spore anche nei basidiomiceti si presentano molto variabili, nella forma, nel colore e nelle ornamentazioni. Presentano generalmente una piccola protuberanza ad una delle due estremità (apiculo) che altro non è che l’appendice che teneva ancorata la spora allo sterigma, e possono avere all’altra estremità un poro germinativo, una sorta di forellino da cui avrà origine il nuovo micelio. Anche per le spore dei basidiomiceti un carattere importante per la determinazione è l’amiloidia ossia la reazione positiva al reattivo di Melzer, la cianofilia ovvero quando la parete sporale si tinge di blu a contatto con il reattivo Blu cotone.
I Giunti a fibbia sono la risultanza di un particolare tipo di accrescimento delle ife mediante un diverticolo prodotto dalla cellula apicale. Non sono presenti in tutti i generi o in tutte le specie di un genere e la loro presenza o assenza è spesso decisiva per la determinazione della specie in esame. Si trovano generalmente ai setti delle ife ma anche alla base dei basidi o basidioli. Nei basidiomiceti inoltre riveste particolare importanza, soprattutto per alcuni generi, lo studio della forma e della disposizione delle ife che costituiscono la Pileipellis (rivestimento del cappello), particolarmente importante ad esempio nel genere Russula, e la Caulopellis (rivestimento del gambo), ad esempio nel genere Mycena.
PRINCIPALI REAZIONI E METODI DI PREPARAZIONE DEL VETRINO
AMILOIDIA
su exsiccata:
1) reidratare il frammento da osservare in KOH 5% per circa 1’
2) fare uno o due lavaggi in acqua e asciugare
3) inserire una goccia di Melzer e attendere 1’ ca.
4) lavare con Cloralio Idrato e asciugare
5) osservare in acqua o in Cloralio Idrato oppure in L4 Tampone alla glicerina
su materiale fresco:
1) escludere il passaggio in KOH 5%
2) procedere come da punto 3
CIANOFILIA
su exsiccata:
1) reidratare per circa 1’ in KOH 5%
2) lavare ed asciugare il tutto per 1-2 volte in acqua
3) inserire una goccia di Blu lattico
4) riscaldare il preparato a fuoco moderato per alcuni secondi, se necessario aggiungere altro Blu lattico
5) togliere il frammento e passarlo su un altro vetrino portaoggetti
6) aggiungere Cloralio Idrato
7) osservare
NOTA: per alcuni generi quali Galerina, Rhodocybe o Lepista non occorre riscaldare il preparato.
CARMINOFILIA O SIDEROFILIA
su exsiccata:
1) reidratare il frammento in KOH 5% per circa 1’
2) lavare ed asciugare il preparato per 1-2 volte in acqua
3) mettere Carmino Acetico
4) riscaldare il tutto a fuoco molto lento agitando con un ferro (non acciaio) per qualche secondo
5) passare il preparato su un altro vetrino portaoggetti e aggiungere Carminio Acetico o Cloralio Idrato
6) schiacciare molto forte il preparato e osservare.
PRINCIPALI REAGENTI E PREPARATI CHIMICI PER L’OSSERVAZIONE MICROSCOPICA
Tavolo di lavoro © Pierluigi Angeli
AMMONIACA (NH3) : si utilizza per la reidratazione di campioni secchi (osservazione delle exsiccata); normalmente è in soluzione acquosa concentrata al 2-3 o 6%.
IDRATO DI POTASSIO (KOH) : ha le stesse proprietà dell’ammoniaca, si usa quindi in exsiccata ad una concentrazione massima del 5%.
IDRATO DI SODIO (NaOH) : identico ai precedenti, anche questo va usato ad una concentrazione non superiore al 5-10%.
IDRATO DI CLORALIO (C Cl3 CH(OH)2) : ha la caretteristica di schiarire.
Nel caso si voglia osservare a lungo il preparato bisogna usare liquidi che eliminano la possibilità di riproduzione dei microbi all’interno del vetrino, il LATTOFENOLO DI AMMAN (soluzione di Amman) composto da Fenolo, Acido Lattico e Glicerina in soluzione acquosa al 20%, è un liquido battericida molto indicato, specialmente se si vogliono fare lunghe osservazioni eseguendo fotomicrografiche; è importante fissare bene il vetrino coprioggetto al vetrino portaoggetti (lutaggio: si ottiene ponendo sopra il vetrino coprioggetto un piccolo peso e sigillando i bordi con dello smalto per unghia), in modo da impedire eventuali fuoriuscite di liquido o infiltrazioni di aria.
Le soluzioni ammoniacali e la potassa si usano anche per l’osservazione di materiale fresco, infatti in alcuni generi (Galerina, Inocybe, Cortinarius) colorano le pareti delle cellule marginali (cistici) provocando una reazione importante dal punto di vista tassonomico.
ROSSO CONGO : è il colorante più usato in Micologia, colora le pareti delle cellule, si adatta bene in molte strutture cellulari; se usato in soluzione ammoniacale è ottimo anche per lavorare con exsiccata.
FLOXINA : ha le stesse caratteristiche del Rosso Congo, colora il citoplasma, è maggiormente usata dai micologi americani, anch’essa si trova in soluzione acquosa che ammoniacale.
BLU COTONE o BLU DI METILE : il più adatto è il Blu Cotone in Lattofenolo, si usa anche come reagente per individuare la cianofilia sporale, ossia la membrana sporale assume una colorazione intensa (genere Entoloma) oppure non si colora (genere Tricholoma).
REAGENTE DI MELZER : si ottiene mescolando una soluzione di Iodio al Cloralio Idrato; serve principalmente a verificare l’amiloidia delle spore. La reazione può essere amiloide (membrana sporale grigio-nerastra o bluastra, apice degli aschi verde-blu) destrinoide o pseudoamiloide (la spora assume una colorazione rossastra), inamiloide.
CARMINIO ACETICO : si usa per osservare una particolare reazione che avviene all’interno dei basidi delle specie appartenenti al gruppo del Lyophyllum s.l. Le granulazioni di questi basidi si definiscono carminofile o siderofile e assumono una colorazione tipicamente grigio-nerastra.
NEO-FUCSINA BASICA (reattivo di ZIEHL) : produce una reazione sulle incrostazioni ifali nella cuticola di alcune Russula , assumendo un importante valore tassonomico.
BLU DI CRESILE : produce una reazione detta metacromatica nelle ife di alcune specie del genere Mycena. La reazione si manifesta facendo diventare la parete ifale esterna di un vivace rosso-vilaceo.
SULFOVANILLINA : mette in evidenza le ife laticifere e i cistidi nei generi Russula e Lactarius.
SOLFATO FERROSO (FeSO4) : si utilizza come reattivo macrochimico nel genere Russula.
TINTURA DI GUAIACO : anche questo è un reattivo macrochimico molto usato nel genere Russula.
MISURAZIONI: COSA, COME E QUANTO
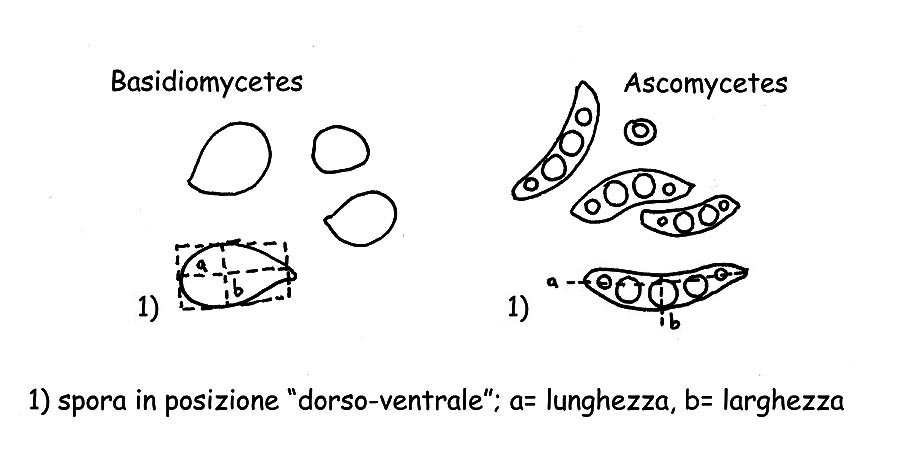
Misurazione spore © Maria Tullii
Spore : Per la misurazione delle spore si usa normalmente rilevare la lunghezza e la larghezza, immaginando una ipotetica forma geometrica (un quadrato, un rettangolo o un cerchio), escludendo le ornamentazioni e l’appendice ilare. Devono essere misurate le spore quando si presentano nella posizione “dorsoventrale”, come da figura (A), normalmente si effettuano circa 32 misurazioni di spore di carpofori maturi, meglio sarebbe effettuare la misurazione su sporata. Per una corretta e valente esposizione delle misure effettuate occorre calcolare sia i “Q” e “QM” (ossia il quoziente ed il quoziente medio) sia il Volume “V” e volume medio “VM” delle spore.
Per il “QM”:
si eliminano le misurazioni marcatamente anomale, sia per quelle troppo piccole sia per quelle troppo grandi, poi di ogni misurazione si troverà il quoziente, (es. 10 × 6,5 – 10,5 × 7 – 9,5 × 7 ecc. 10/6,5 = 1,53846153 che si arrotonderà a 1,53 + 1,5 + 1,35 ecc. = 4,38, dividendo poi per il numero delle misurazioni di cui è stato trovato il quoziente si otterrà il “QM”, in questo esempio 4,38/3 = QM 1,46;
per il “Q”:
si riporterà, dai quozienti ottenuti per il “QM” il più piccolo ed il più grande come per indicare la misurazione delle spore Q= 1,35 – 1,53.
Per quanto riguarda il volume delle spore si prosegue come segue:
si rileverà la spora più piccola del nostro fungo, di essa si calcolerà il volume con la seguente formula V= 4π/3 × (a/2)² × b/2 (a= diametro più piccolo, b= diametro più grande).
Si rileverà la spora più grande e si calcolerà il volume come per la precedente V1= 4π/3 × (A/2)² × B/2 (A= diametro più piccolo B= diametro più grande) si proseguirà poi con la formula VM = (V + V1)/2, ad esempio: in una serie di misurazioni abbiamo la spora più piccola che misura 6,5 × 4 µm e la più grande 8 × 5,5 µm applicando la formula avremo: V = (4×3,14/3) × (4/2)² × 6,5/2 = 54,34; V1 = (4×3,14/3) × (5,5/2)² × 8/2 = 126,40; VM = (54,34 + 126,40) / 2 = VM 90,4.
Basidi e Cistidi : Si effettua la misurazione della lunghezza, partendo dall’attaccatura del basidio o del cistidio al sottoimenio, la larghezza si prende nel punto mediano o normalmente nel punto più largo.

Amanita muscaria – Amanita phalloides – Astraeus hygrometricus

Astraeus hygrometricus – Boletus aestivalis – Boletus edulis – Suillus granulatus

Tricholoma columbetta – Suillus granulatus – Tuber melanosporum – Tuber magnatum – Tuber melanosporum – Macrolepiota procera

Macrolepiota procera – Russula vesca – Otidea onotica

Verpa bohemica – Morchella esculenta – Marasmius oreades – Mutinus elegans – Macrolepiota procera

Leucoagaricus americanus – Amanita citrina – Amanita muscaria

Amanita muscaria – Amanita rubescens – Boletus aereus

Boletus aereus – Laccaria amethystina – Cortinarius rufoolivaceus

Cortinarius rufoolivaceus – Gyromitra infula – Cortinarius lividoviolaceus

Cortinarius lividoviolaceus – Amanita spissa


Amanita spissa – Cantharellus cibarius – Amanita mairei

Amanita mairei – Clitocybe gibba

Calvatia excipuliformis – Calvatia craniiformis – Clavariadelphus pistillaris

Clavariadelphus pistillaris – Clitocybe nebularis

Cortinarius turmalis – Clavulina cinerea – Gomphidius glutinosus

Gomphidius glutinosus – Clitocybe odora

Russula atropurpurea – Calvatia utriformis – Crucibulum laeve

Lactarius deliciosus – Suillus grevillei

Suillus grevillei – Russula mairei – Leccinum aurantiacum

Leccinum aurantiacum – Mycena inclinata – Oudemansiella mucida

Oudemansiella mucida – Mycena renatii – Lepiota clypeolaria – Lactarius torminosus

Lactarius torminosus – Badhamia utricularis

Badhamia utricularis

Clavulina rugosa

Russula foetens – Psathyrella caput-medusae – Cantharellus lutescens – Suillus luteus

Suillus luteus – Amanita muscaria – Stropharia aeruginosa – Tricholoma saponaceum


Amanita pantherina – Aleuria aurantia

Boletus aestivalis – Entoloma lividoalbum

Boletus edulis – Boletus aestivalis

Chroogomphus rutilus – Calocera viscosa – Gloeophyllum sepiarium

Stereum hirsutum – Lactarius aurantiacus – Fuligo septica

Fuligo septica

Fuligo septica – Cortinarius variecolor – Fistulina hepatica

Fistulina hepatica – Galerina marginata – Hypholoma sublateritium

Hypholoma sublateritium – Hygrophoropsis aurantiaca – Hypholoma fasciculare

Hypholoma fasciculare – Ischnoderma benzoinum

Ischnoderma benzoinum – Leucocortinarius bulbiger – Leccinum holopus – Spathularia flavida

Spathularia flavida – Paxillus involutus

Clitocybe costata – Fomitopsis pinicola – Lycoperdon perlatum

Lycoperdon perlatum – Lepista flaccida
= Lepista inversa – Suillus grevillei – Russula integra

Russula integra – Lactarius intermedius – Oudemansiella radicata

Oudemansiella radicata – Suillus granulatus – Clathrus ruber

Clathrus ruber – Tricholoma acerbum – Russula xerampelina

Ramaria lacteobrunnescens – Armillaria mellea = Armillariella mellea


Amanita vaginata – Amanita phalloides

Clitocybe costata – Boletus aestivalis – Boletus regius


Boletus edulis – Agaricus silvicola – Clitocybe candicans – Amanita rubescens

Cantharellus cibarius – Chroogomphus rutilus

Calocera viscosa – Clavulina rugosa

Cortinarius anomalus – Cortinarius cliducus – Cortinarius infractus

Cortinarius olivascentium – Cystolepiota seminuda

Cribraria argillacea – Ceratiomyxa fruticulosa

Ceratiomyxa fruticulosa = Famintzia fruticulosa

Ceratiomyxa fruticulosa var. porioides

Clavulina coralloides

Lactarius piperatus – Coprinus micaceus

Collybia fusipes – Cortinarius cotoneus – Dacrymyces abietinum

Ife miceliari

Ife miceliari – Hemimycena gracilis – Hemimycena pseudolactea


Hypholoma fasciculare – Hebeloma edurum

Lycoperdon perlatum – Lactarius deliciosus

Lactarius salmonicolor – Lactarius volemus

Lactarius porninsis – Lactarius serifluus – Leccinum aurantiacum

Lycogala epidendron

Lycogala epidendron

Lactarius circellatus – Lactarius intermedius – Laetiporus sulphureus – Hypholoma sublateritium – Leccinum carpini

Leucopaxillus candidus – Macrolepiota mastoidea

Lycoperdon echinatum – Lycoperdon molle

Macrolepiota procera – Mycena pura

Phaeomarasmius erinaceus – Merulius tremellosus

Phellodon tomentosus – Pluteus cervinus

Polyporus brumalis – Paxillus involutus

Russula cyanoxantha – Russula risigallina

Russula virescens – Russula aurea

Russula integra – Russula velutipes = Russula aurora

Russula delica – Marasmius oreades – Marasmius bulliardii – Scleroderma citrinum

Sowerbyella imperialis

Stropharia caerulea – Suillus granulatus – Trametes versicolor – Penicillum digitatum

Stemonitis axifera : fase giovanile

Stemonitis axifera : giovanile, in crescita e adulta
Stemonitis axifera adulta

Xerocomus subtomentosus – Xerocomus chrysenteron – Leccinum scabrum

Amanita rubescens – Amanita lividopallescens
Boletus satanas

Boletus satanas – Pholiota squarrosa
Ramaria formosa
Ramaria formosa – Tricholoma portentosum

Boletus edulis in età matura – Crucibulum laeve

Lasiochlaena benzoina = Ischnoderma benzoinum – Lycoperdon echinatum

Trametes versicolor – Trametes gibbosa

Phaeolus schweitnizii – Coprinus niveus

Cortinarius herculeus – Kretzschmaria deusta = Ustulina deusta : la forma grigia è l’ Anamorfo, quella nera il Teleomorfo

Inocybe geophylla – Tricholoma terreum – Perenniporia narymica – Laccaria bicolor

Stropharia caerulea – Coprinus comatus

Coprinus comatus – Amanita caesarea – Spore di Tuber melanosporum

Spore di Boletus edulis – Otidea umbrina – Tuber magnatum
© Giuseppe Mazza