Phylum : Amoebozoa

Testo © Prof. Giorgio Venturini

Amoeba proteus in colorazione vitale, col Rosso Neutro che si concentra nei lisosomi © Giuseppe Mazza
Amoeba proteus Pallas, 1766, è la specie più nota di un genere di protisti precedentemente inserito nel phylum Rhizopoda, ma attualmente, sulla base di dati molecolari, classificato come phylum Amoebozoa, appartenente al Regno dei protisti. Si deve comunque ricordare che la classificazione dei protisti è attualmente molto discussa.
La maggior parte delle specie di ameba sono a vita libera nel suolo, nel fango o nell’acqua, dove si cibano di batteri, piccoli protisti e altri organismi unicellulari; soltanto poche specie sono parassite.
Amoeba proteus è la specie meglio studiata: si tratta di un organismo unicellulare di circa 250 micron (μm) di diametro, che può raggiungere anche i 400-500 micron, capace di emettere prolungamenti citoplasmatici a forma di lobi, gli pseudopodi, utilizzati per la locomozione e la cattura delle prede. Il genere Chaos, affine ad Amoeba, comprende le più grandi amebe conosciute, tra cui ricordiamo Chaos carolinense che può raggiungere i 5 mm.
Osservata al microscopio a fresco e senza colorazioni l’ameba ha l’aspetto di una massarella di forma irregolare, priva di colorazione e traslucida, animata da continui movimenti che consistono in cambiamento di forma, emissione di pseudopodi e continuo fluire del materiale granuloso citoplasmatico. Al suo interno oltre a numerosi granuli si osserva un nucleo ed un grosso vacuolo tondeggiante. Al contrario del genere Chaos i cui membri sono plurinucleati, Amoeba proteus è uninucleata.
L’habitat dell’ameba consiste essenzialmente nella melma sul fondo di pozze d’acqua dolce, laghi e corsi d’acqua con scarsa corrente, spesso sotto la vegetazione acquatica, e anche nei suoli molto umidi.
Il nome del genere Amoeba, ameba in italiano, deriva dal greco “amoibé” (ἀμοιβή) = cambiamento e si riferisce al suo mutare di forma; Proteo nella mitologia Greca è il nome di una divinità marina, detto anche “il vecchio del mare”, capace di trasformarsi assumendo l’aspetto di qualunque animale o pianta, ma anche di oggetti come acqua o fuoco.
La più antica descrizione di un’ameba è dovuta a August Johann Rösel von Rosenhof nobile austriaco, pittore di miniature, naturalista ed entomologo che nel 1755 la battezzò “Der Kleine Proteus” (“il piccolo Proteo”). Grazie alla sua abilità di pittore questo studioso ci ha lasciato bellissime immagini dell’ameba e di moltissimi altri organismi.
Storicamente i microscopisti hanno suddiviso il citoplasma dell’ameba in due parti, un “ectoplasma” esterno poco granuloso e a contatto con la membrana cellulare e un “endoplasma” granulare interno a sua volta distinguibile in una zona più viscosa, detta plasmagel ed una meno viscosa, detta plasmasol. In realtà queste suddivisioni rispecchiano condizioni funzionali diverse del citoplasma e sono continuamente variabili.
Locomozione
Per comprendere la locomozione di un’ameba possiamo pensare al nostro passo, che prevede la proiezione in avanti di una gamba e il successivo contatto del piede con il suolo. Il piede ora ben poggiato a terra serve come ancoraggio per tirare in avanti il corpo grazie alla contrazione dei muscoli dell’anca mentre l’altro piede, prima fisso al suolo, si solleva.
Anche nell’ameba il principio è lo stesso: proiezione in avanti di uno pseudopodio, ancoraggio di questo al substrato e trazione del corpo cellulare in avanti, mentre i precedenti ancoraggi si distaccano.
Si tratta ora di capire in che modo si formi uno pseudopodio, come si allunghi e si ancori al substrato e come si verifichi la trazione in avanti del corpo cellulare.
Per spiegare i movimenti dell’ameba e di altre cellule dotate di movimenti ameboidi sono state proposte nel tempo diverse teorie che in genere invocano i cambiamenti di consistenza delle diverse parti del citoplasma che passano alternativamente da uno stato di sol ad uno di gel (cioè da liquido a gelatinoso e viceversa), permettendo una sorta di spremitura del citoplasma verso il fronte di avanzamento. Non appare chiaro in queste teorie quali siano i meccanismi che provocano questi cambiamenti di consistenza, come vengano diretti, come in realtà generino un movimento e quali siano le fonti di energia per compiere questo lavoro.
Più recentemente è stato preso in esame il ruolo di alcune proteine presenti nel citoplasma dell’ameba e ben note negli organismi pluricellulari, che formano un vero e proprio scheletro flessibile intracellulare (citoscheletro), capaci di allungarsi e accorciarsi attivamente oltre che di funzionare come rotaie su cui possono muoversi altre proteine, veri e propri motori molecolari capaci trascinare a loro volta altri filamenti proteici. Queste proteine citoscheletriche e i relativi motori molecolari sono anche alla base del movimento dei nostri muscoli.
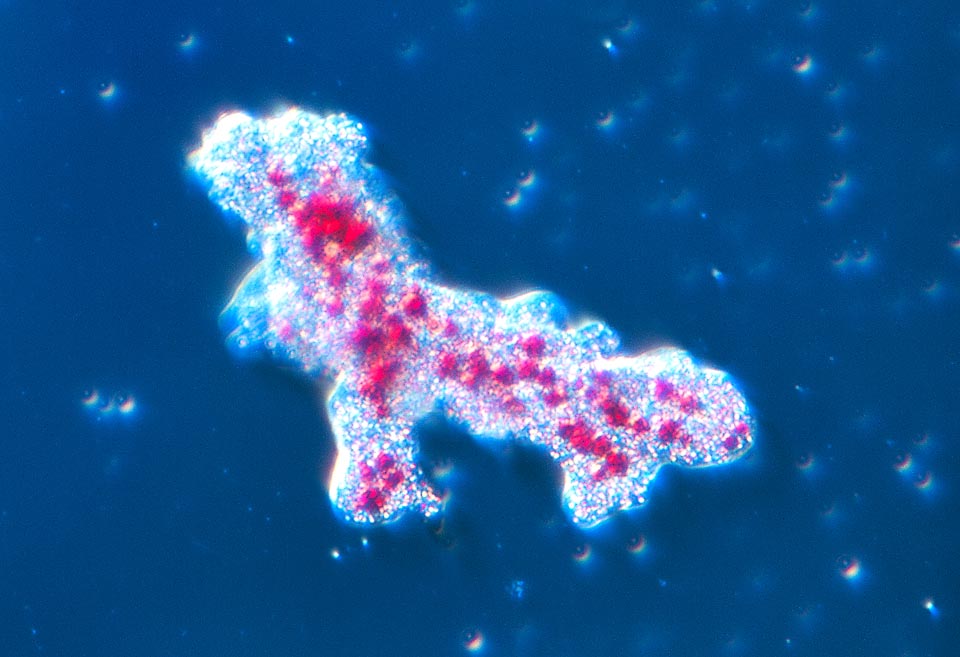
L’habitat dell’ameba consiste essenzialmente nella melma sul fondo di pozze d’acqua dolce, laghi e corsi d’acqua con scarsa corrente di tutto il mondo © Giuseppe Mazza
Cerchiamo qui di delineare un possibile meccanismo (che comunque non esclude il possibile intervento di altri fenomeni).
La proteina principale che forma i filamenti citoscheletrici è detta actina ed è una molecola di forma approssimativamente globulare, abbondante nel citoplasma dell’ameba come in quello delle nostre cellule. Le molecole di actina possono aggregarsi reversibilmente per formare un filamento, come una specie di collana di perle, che può allungarsi o accorciarsi per aggiunta o sottrazione di molecole globulari. Si pensa che la protrusione degli pseudopodi possa essere almeno in parte generata dall’allungamento dei filamenti di actina che, associati in fasci paralleli, verrebbero a spingere in avanti la membrana.
In aggiunta o in alternativa a questa spiegazione si è invocato un ruolo della actina e delle proteine motrici ad essa associate nel produrre una forza contrattile capace di indurre per pressione dei flussi di citoplasma verso il fronte di avanzamento.
Anche il cambiamento della consistenza del citoplasma da uno stato più liquido ad uno più solido è conseguenza della aggregazione delle molecole di actina. Altre proteine interagiscono con la actina formando dei legami crociati tra i filamenti e generando quindi una rete tridimensionale.
Per comprendere meglio questo fenomeno possiamo pensare a quello che avviene quando il brodo di carne raffreddandosi si trasforma in gelatina: le molecole proteiche del collagene, separate l’una dall’altra nel brodo bollente, si aggregano a formare dei filamenti che creano una rete tridimensionale che rassoda la gelatina. Nell’esempio della gelatina si forma una rete con fili a orientamento casuale, mentre nei nostri tessuti, grazie all’intervento delle cellule, il collagene si aggrega a formare dei robusti fasci paralleli, ad esempio i tendini. Quando lo pseudopodio si è allungato delle proteine della membrana formano degli ancoraggi al substrato.
Quali sono i fattori che stimolano la formazione dei filamenti di actina? Cosa è per una ameba “anteriore” o “posteriore” ?
Sappiamo che le amebe sono sensibili a segnali presenti nell’ambiente (tropismi positivi o negativi), ad esempio vengono attratte da molecole rilasciate da fonti di cibo: queste molecole si legano a proteine recettoriali presenti sulla membrana dell’ameba e attivano una risposta molecolare che stimola la aggregazione della actina globulare per formare i filamenti. È interessante notare che le molecole che stimolano la aggregazione della actina nell’ameba sono le stesse che svolgono lo stesso compito nei nostri globuli bianchi o in molte altre cellule mobili. È quindi il segnale chimico proveniente dall’ambiente che stimola la protrusione degli pseudopodi e che determina quale sarà il polo anteriore della cellula. Quando il segnale scompare gli pseudopodi cessano di allungarsi in quella direzione dal momento che i filamenti di actina si disaggregano. La tappa successiva richiede il trascinamento o la spinta del corpo cellulare in direzione dello pseudopodio: qui entrano in gioco le proteine motrici (miosine) che come dei veri e propri motori molecolari “camminano” sui filamenti di actina e determinano il movimento delle impalcature citoscheletriche del citoplasma in direzione dell’estremità avanzante, mentre i vecchi ancoraggi proteici al substrato si distaccano.
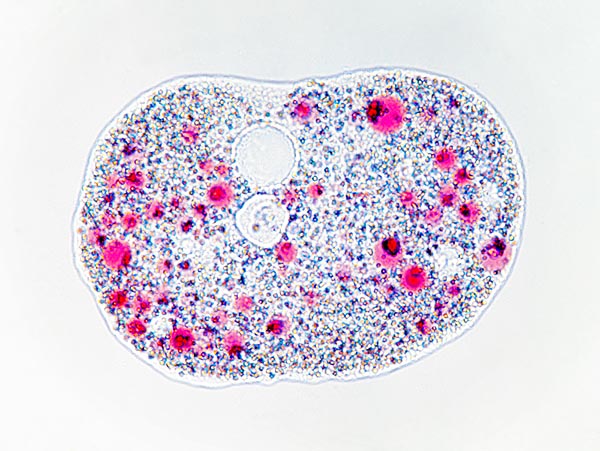
Amoeba proteus a riposo. Al centro in alto si nota il vacuolo © Giuseppe Mazza
Questa interazione tra actina e miosine è analoga a quella che opera nei nostri muscoli.
Naturalmente sia l’allungamento dei filamenti per apposizione di nuove sub unità che l’attività dei motori molecolari come la miosina richiedono energia, che viene fornita, come anche per la maggior parte dei fenomeni cellulari richiedenti energia, dalla ben nota molecola di ATP, molecola ricca di energia continuamente prodotta dal metabolismo di tutte le cellule. Anche frammenti di citoplasma isolati dal corpo cellulare, in presenza di ATP esibiscono movimenti.
Una conferma del ruolo essenziale dei filamenti di actina nella locomozione dell’ameba proviene da esperimenti in cui alle amebe è stato somministrato un farmaco (citocalasina) che impedisce la formazione dei filamenti di actina. In queste condizioni l’ameba non è più in grado di emettere pseudopodi e di spostarsi.
Può stupire il fatto che gli stessi macchinari molecolari operino nei movimenti dell’ameba, nei nostri globuli bianchi o nei nostri muscoli: possiamo spiegarci queste similitudini riflettendo sul fatto che i meccanismi del movimento cellulare si sono evoluti nel lontano progenitore che, quasi un miliardo di anni or sono, ha dato origine alla linea evolutiva dei protozoi e a quella che condurrà agli esseri umani. Dunque l’actina dell’ameba è identica a quella umana? Certamente non identica, dal momento che l’actina del progenitore comune ha accumulato cambiamenti evolutivi (mutazioni) per quasi un miliardo di anni, sia evolvendosi verso la actina dell’ameba moderna che verso quella del moderno uomo, ma le sue caratteristiche funzionali di base sono rimaste uguali.
Alimentazione
L’ameba è un organismo predatore che si nutre inglobando mediante fagocitosi altri protisti o batteri. Il fenomeno è reso possibile dalla sua capacità di percepire la presenza delle prede grazie a segnali chimici o di contatto e soprattutto di emettere pseudopodi mediante i quali circonda il microorganismo e lo ingloba in una vescicola di membrana (vacuolo di fagocitosi o fagosoma). In realtà per una rappresentazione più corretta del fenomeno dobbiamo tener presente la sua tridimensionalità e pensare in questo caso agli pseudopodi come a delle labbra che si chiudono intorno alla preda per inserirla nella “bocca”, cioè nel fagosoma.
È intuitivo che la membrana che circonda il fagosoma deriva dalla membrana cellulare e quindi ogni fagocitosi sottrae materiale all’involucro. Se consideriamo che una ameba in una giornata esegue probabilmente una media di 100 fagocitosi, in ognuna delle quali sottrae circa il 10% alla propria superficie di membrana, possiamo calcolare che ogni giorno venga sottratto l’equivalente di 10 superfici cellulari. Questo è reso possibile da concomitanti fenomeni di esocitosi che restituiscono alla superficie la membrana dei vecchi fagosomi o membrana neo-formata.
Per comprendere correttamente questo continuo riciclo di membrane in un cellula e il trasferimento del contenuto delle vescicole dobbiamo pensare che membrana è una pellicola fluida e il comportamento di una cellula e dei suoi sistemi di vescicole può essere paragonato a quello di gocce d’olio poste in un piatto pieno d’acqua: le gocce possono fondersi, unendo i loro contenuti o venir frammentate per poi fondersi ancora. Allo stesso modo dalla membrana della cellula possono staccarsi vescicole pronte a fondersi di nuovo, trasferendo il loro contenuto e la loro membrana in un continuo interscambio.
Dopo la fagocitosi il materiale inglobato viene ora avviato alla digestione, facendo fondere il fagosoma con altre vescicole membranose contenenti enzimi digestivi, equivalenti funzionalmente a quelli presenti nel nostro apparato digerente. Dal momento che questi enzimi funzionano correttamente soltanto in ambiente molto acido, delle pompe molecolari presenti sulla membrana del fagosoma accumulano al suo interno ioni idrogeno (acidità corrisponde ad alta concentrazione di ioni idrogeno). La vescicola risultante, acida e contenente il materiale fagocitato e gli enzimi digestivi, prende il nome di lisosoma e svolge la funzione di apparato digerente cellulare.
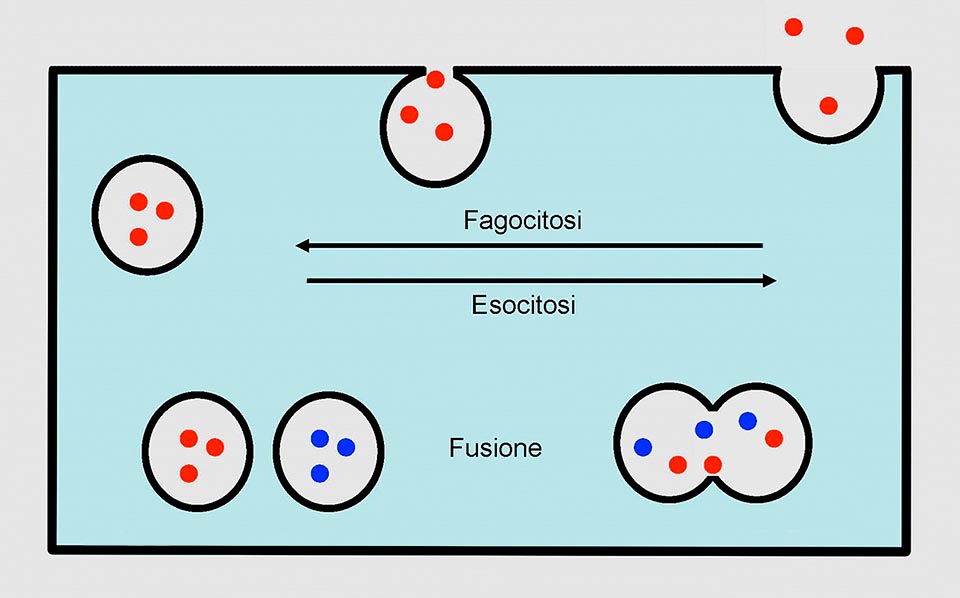
Nell’ameba la membrana con la fagocitosi produce una vescicola che si fonde con il lisosoma, dopo la digestione i prodotti di scarto vengono espulsi con una esocitosi che inoltre restituisce alla membrana la superficie sottratta con la fagocitosi. Anche il vacuolo contrattile espelle il suo contenuto mediante una esocitosi © Giorgio Venturini
Quando gli enzimi avranno completato la loro azione il lisosoma conterrà materiale digerito, utile per l’ameba, oltre a prodotti di scarto e materiale indigeribile. A questo punto delle proteine trasportatrici presenti sulla membrana del lisosma trasferiscono nel citoplasma le sostanze utili (ad esempio zuccheri o amminoacidi derivanti dalla digestione). Infine il lisosoma si avvicina alla membrana cellulare e si fonde con essa, riversando all’esterno i prodotti di scarto. In ogni caso non si verifica mai la rottura del lisosoma e il riversamento di tutto il suo contenuto nel citoplasma: se ciò si dovesse verificare la cellula si auto-digerirebbe!
È facile il confronto con la nostra alimentazione: il cibo ingerito viene digerito dagli enzimi presenti nello stomaco e nell’intestino, le sostanze utili ottenute dalla digestione vengono assorbite dalla parete intestinale e trasferite al sangue mentre i prodotti di scarto vengono espulsi con le feci. La rottura della parete dell’intestino e dello stomaco (perforazione) ha conseguenze gravissime.
Mentre la fagocitosi permette all’ameba di inglobare strutture voluminose come batteri o protisti, questo organismo può assumere dall’ambiente sostanze nutrienti mediante un processo detto pinocitosi (dal greco “πινω” (pino), che significa bevo), che comunque si basa su principi simili alla fagocitosi, in quanto permette di inserire in vescicole membranose acqua e soluti macromolecolari che poi verranno inviati al lisosoma.
Si deve ricordare infine che la membrana cellulare dell’ameba possiede sofisticati meccanismi di permeabilità selettiva che permettono il passaggio di singole molecole di piccole dimensioni, come ad esempio zuccheri, dall’esterno direttamente al citoplasma, senza formazione di vescicole.
Il vacuolo contrattile e la regolazione idrica
Vivendo in ambienti di acqua dolce l’ameba per sopravvivere deve affrontare il problema della regolazione del proprio contenuto di acqua. Il citoplasma infatti può essere considerato come una soluzione molto concentrata di sostanze organiche (come ad esempio proteine o zuccheri) e inorganiche, separato, grazie alla membrana cellulare, da un ambiente esterno acquoso a bassa concentrazione di soluti, e quindi più ricco di acqua. Essendo la membrana cellulare permeabile all’acqua la elevata concentrazione di soluti interna richiama verso l’interno acqua per effetto osmotico. Il risultato inevitabile, in assenza di meccanismi di regolazione, sarebbe quello di far rigonfiare la cellula fino a farla scoppiare.
L’ameba riesce a far fronte a questo problema grazie ad un organello cellulare, il cosiddetto vacuolo contrattile, una vescicola membranosa che accumula al proprio interno l’acqua in eccesso e poi la espelle all’esterno della cellula.
Il meccanismo mediante il quale il vacuolo riesce ad accumulare l’acqua è rimasto a lungo oscuro, il suo recente chiarimento ha permesso di svelare anche altre funzioni di questa struttura.
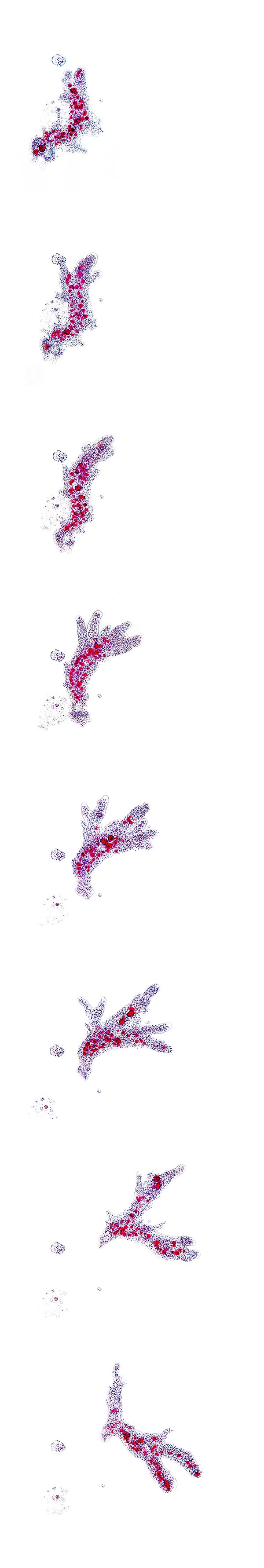
Sequenza del movimento di una Amoeba proteus © Giuseppe Mazza
La membrana del vacuolo è dotata di canali permeabili all’acqua e di pompe molecolari capaci di trasferire attivamente al suo interno diverse sostanze, tra cui soprattutto ioni idrogeno. Grazie a questo trasporto l’interno del vacuolo presenta una elevatissima concentrazione di ioni che, per effetto osmotico, attrae acqua dal citoplasma. In questo modo l’acqua, che per la azione osmotica del citoplasma era entrata nella cellula, per effetto osmotico si accumula nel vacuolo (se consideriamo il gradiente della concentrazione dei soluti, questa risulta minima nell’ambiente esterno, intermedia nel citoplasma e massima nel vacuolo, quindi l’acqua si trasferirà dall’esterno al citoplasma e da questo al vacuolo).
A questo punto il vacuolo si sposta fino a giungere a contatto con la membrana cellulare e si apre verso l’esterno riversando il suo contenuto di acqua e di soluti. In realtà sulla membrana del vacuolo sono presenti, oltre alle pompe per gli ioni idrogeno, anche altri trasportatori che accumulano al suo interno diverse sostanze, come ad esempio scarti metabolici quali anidride carbonica, urea o acido urico, che verranno espulsi all’esterno quando il vacuolo riversa il suo contenuto fuori della cellula. Di conseguenza il vacuolo non funziona soltanto come organulo regolatore del contenuto di acqua ma anche come organulo escretore che elimina i rifiuti del metabolismo. Per espellere il suo contenuto il vacuolo fonde la sua membrana con la membrana cellulare e quindi a questo punto il vacuolo scompare: un nuovo vacuolo si formerà ex novo mediante la produzione di una nuova vescicola.
Come ovvio il trasporto di ioni idrogeno e di altre sostanze all’interno del vacuolo richiede energia, che viene fornita dalla molecola di ATP.
Il termine vacuolo contrattile suggerisce una sua capacità di contrarsi attivamente, suggerita dalle osservazioni dei microscopisti che notavano suoi periodici cambiamenti di volume. Sembra in realtà che questi cambiamenti siano essenzialmente dovuti all’accumulo di acqua e non a fenomeni di contrazione attiva.
Nelle amebe che vivono in acqua di mare, che non hanno il problema del rigonfiamento visto che l’acqua di mare ha concentrazione salina superiore a quella del citoplasma, il vacuolo non esiste (queste amebe avranno invece il problema opposto della disidratazione).
Genetica e riproduzione
Come nel caso di altre amebe giganti anche Amoeba proteus ha nel suo nucleo un grandissimo numero di cromosomi, circa 250 coppie, ed una quantità enorme di DNA, stimata in circa 290 miliardi di basi (nell’uomo abbiamo 23 coppie di cromosomi e 3 miliardi di basi). Gli altissimi valori riscontrati in Amoeba proteus derivano da fenomeni di poliploidia, cioè i cromosomi si sono ripetutamente duplicati in copie identiche. I numeri citati sono inoltre variabili durante il ciclo vitale del protozoo. Non sono chiari i significati funzionali di questi fenomeni.
Riproduzione
La riproduzione dell’ Amoeba proteus è un processo periodico che si ripete a intervalli che dipendono dal tasso di accrescimento. Un’ameba inizierà a dividersi quando raggiunge una dimensione sufficiente, ad esempio 0,2-0,3 mm. La riproduzione è asessuata e avviene in genere per scissione binaria. Prima di dividersi la cellula retrae gli pseudopodi e assume una forma tondeggiante, quindi si verifica una divisione nucleare mitotica seguita da suddivisione del citoplasma che avviene grazie ad una strozzatura del citoplasma (dovuta alle stesse proteine contrattili che agiscono nella locomozione) seguita dalla attività degli pseudopodi che trascinano in direzioni opposte le due cellule figlie. Il processo dura in genere tra 30 e 60 minuti.
Sono stati descritti fenomeni di fissione multipla, che avverrebbe in condizioni sfavorevoli di scarsa alimentazione o di disidratazione: l’ameba si arrotonda e si ricopre di una cisti resistente e impermeabile, che le permette di sopravvivere fino a quando le condizioni divengano favorevoli. A questo punto una divisione nucleare multipla produce numerosissimi nuclei che si raccolgono alla periferia della cellula e infine si separano in tante nuove cellule che fuoriescono quando la cisti si idrata e scoppia.
L’incistamento sembra poter avvenire anche indipendentemente dalla moltiplicazione e rappresenta quindi un sistema per superare periodi di condizioni avverse, come l’essiccamento della pozza d’acqua.
Alcuni microscopisti hanno descritto fenomeni che fanno pensare ad una coniugazione, ma il significato di queste osservazioni non è chiaro.
Simbiosi e parassitismo
Anche se Ameba proteus è in grado di fagocitare e digerire la maggior parte dei batteri, esiste almeno una specie di batterio, simile alla Legionella, l’agente patogeno della malattia dei legionari, che è in grado di sopravvivere all’interno del fagosoma, evitando di essere digerito. Questo batterio stabilisce una relazione simbiotica con l’ameba che induce in ambedue le specie dei cambiamenti che le rendono mutuamente dipendenti.
In alcune specie di ameba sono state riscontrate infezioni da parte di virus caratterizzati da dimensioni eccezionalmente grandi, come i Mimi virus o il virus Pandora. Questi virus hanno diametri tra 400 e 1000 nanometri (mn, miliardesimi di metro) e il loro DNA ammonta a uno o due milioni di basi. Consideriamo per confronto che la maggior parte dei virus hanno dimensioni di poche decine di nm e il loro DNA è di poche decine di migliaia di basi.
Predatori dell’ameba
Le amebe sono oggetto di predazione da parte di diversi organismi, tra cui nematodi, piccoli pesci, crostacei e molluschi, da cui possono difendersi mediante la secrezione di sostanze tossiche o repellenti.
Amebe patogene per l’uomo
La patogenicità delle amebe è un argomento non completamente chiarito, dal momento che soltanto pochi ceppi delle specie potenzialmente implicate risultano permanentemente patogeni per l’uomo e nella maggior parte dei casi perdono facilmente la loro infettività.
I generi più importanti sono Naegleria e Balamuthia, che possono causare gravi encefaliti, Acanthamoeba, causa di meningo-encefatite e di cheratite, o Hartmannella, cui sono imputate delle cheratiti.
Oltre a questo si deve ricordare che alcune amebe possono essere infettate da batteri patogeni per l’uomo e quindi fungere da vettori di malattia.
La specie sicuramente patogena più importante è però Entamoeba histolytica, agente della amebiasi intestinale, una infezione intestinale che può essere molto grave, con complicanze anche mortali. Si stima che 50 milioni di persone nel mondo siano infestate da questa ameba, soprattutto nei paesi tropicali, con 50000-100000 morti per anno. Nella maggior parte dei casi l’infestazione è asintomatica, ma in circa il 10% dei casi provoca forme di dissenteria.
Le amebe invadono la mucosa intestinale e se ne nutrono, producendo ulcerazioni, con perdita di sangue che può provocare gravi anemie e possibili perforazioni dagli esiti gravissimi. Le ulcerazioni possono permettere ai protozoi di raggiungere il circolo sanguigno e quindi di essere trasportati ad organi come il fegato o il cervello provocando gravi ascessi anche mortali.
Quando raggiungono il colon, in seguito alla progressiva disidratazione del contenuto intestinale, le amebe producono una cisti che verrà emessa con le feci e che rappresenta la forma capace di infettare altri individui. La cisti matura presenta quattro nuclei e proprio la sua ricerca mediante l’esame microscopico delle feci rappresenta il classico metodo diagnostico. La cisti è estremamente resistente e può sopravvivere a lungo nell’acqua o nel terreno.
Il contagio avviene per ingestione di acqua o cibo contaminati dalle feci contenenti le cisti che, quando raggiungono l’intestino del nuovo ospite si trasformano nel trofozoite, cioè nell’ameba attiva capace di moltiplicarsi e di invadere la mucosa.
L’amebiasi, è un esempio tipico di infezione a contagio fecale, descritto dagli inglesi come “Ciclo F”: feces (feci), fingers (dita), flies (mosche), food (cibi). I parassiti presenti nelle feci, tramite le mani o le mosche giungono ai cibi. Si riportano numerosi aneddoti su casi di contagio multiplo, tra cui quello di un cuoco di una nave da crociera che, evidentemente poco attento all’igiene delle sue mani, avrebbe contagiato tutti i passeggeri! (Ricordiamo che nei paesi anglosassoni nei gabinetti pubblici sono affissi cartelli “Now wash your hands!”)
Al genere Entamoeba appartengono numerose specie, alcune delle quali, come Entamoeba histolytica, Entamoeba dispar, Entamoeba moshkovskii, Entamoeba polecki, Entamoeba coli, e Entamoeba hartmanni possono riesiedere nell’intestino umano. Soltanto Entamoeba histolytica è sicuramente patogena. La diagnosi differenziale si basa sulle caratteristche della cisti, tetranucleata in Entamoeba histolytica, e soprattutto su metodiche immunologiche o di biologia molecolare.
L’ameba nella cultura popolare
Nella canzone “A very cellular song” del gruppo musicale inglese “The incredible string band” si descrive la vita dal punto di vista di un’ameba: “Amoebas are very small, There’s absolutely no strife Living the timeless life, I don’t need a wife Living the timeless life. If I need a friend, I just give a wriggle split right down the middle and when I look there’s two of me both as handsome as can be. Oh, here we go slithering, here we go slithering and squelching on …”
“Le amebe sono molto piccole, non c’è assolutamente alcuna lotta vivendo una vita senza tempo, non mi serve una moglie, se ho bisogno di un amico faccio un guizzo e mi divido a metà e se guardo vedo due di me, tutti e due belli quanto si può essere. Ce ne andiamo strisciando e spiaccicandoci….”
Sinonimi
Volvox proteus Pallas, 1766; Proteus diffluens O.F. Müller, 1786; Chaos diffluens (O.F. Müller, 1786) Schaeffer, 1926.