Famiglia : Apidae

Testo © DrSc Giuliano Russini – Biologo Zoologo

Un’ Apis mellifera, in fase d’atterraggio su un fiore, con la proboscide distesa © Giuseppe Mazza
Classificazione zoologica:
Tipo: Artropodi (Arthropoda), sottotipo Antennati (Antennata), classe Insetti (Insecta), sottoclasse Pterigoti (Pterygota), ordine Imenotteri (Hymenoptera), sottordine Apocriti (Apocrita), famiglia Apidi (Apidae), genere Apis, specie Ape domestica (Apis mellifera Linnaeus, 1758).
Fin dalla più remota antichità (civiltà Sumera, Egizia, Fenicia, ecc.) è nota la vita sociale delle api. Biologi ed allevatori ci hanno lasciato notizie, spesso ben precise sul loro allevamento (apicoltura), su come avere sempre a disposizione il miele, biondo e dolce frutto dell’operosità di questi speciali insetti.
Ma per una conoscenza precisa e approfondita dei costumi, della metamorfosi e del ciclo biologico complessivo della società delle api, si è dovuto aspettare l’epoca moderna, con la costruzione di arnie artificiali e trasparenti, con cui osservare, in qualsiasi momento, le intricate matasse sociali-interattive di questi imenotteri.
Nonostante ciò, in verità, bisogna riconoscere che molti dei loro misteri non sono ancora stati svelati (questo vale per tutti gli insetti eusociali: quelli cioè in cui è evidente una società organizzata in caste, come vespe, formiche, termiti, ecc,.) o almeno, non si è ancora potuto darne una spiegazione esaustiva.
Nella società delle api, con un’architettura suddivisa in caste, che interagiscono secondo moduli sociobiologici e socio-comportamentali molto raffinati, esistono tre tipi di individui, ciascuno delimitante una casta: femmine feconde o “regine”, femmine sterili od “operaie” e infine i maschi, detti anche “fuchi” o “pecchioni”, che sono in realtà i membri più pigri dell’intera società, dediti solo al piacevole fenomeno della riproduzione.
Tale società, è definita “pluriannuale” per l’enorme numero di generazioni, che caratterizzano la copiosa prole della regina, a tipo “matriarcale” per la presenza della regina a suo comando e “polimorfica”, dato che ad ognuna delle tre caste corrisponde un morfotipo ben preciso.

Col trasporto pollinico, le api permettono la fecondazione incrociata © Mazza
La regina di Apis mellifera o Apis mellifica, come successivamente fu chiamata dallo stesso Linneo, nome ancora oggi utilizzato da alcuni apicoltori, ha dimensioni maggiori degli altri abitanti del favo, raggiungendo anche i 2 cm di lunghezza, con un addome massiccio e allungato, che contiene fino a 180 ovarioli.
Non possiede organi specializzati per la raccolta del polline, né “ghiandole della cera, ciripare”, né “ghiandole sopracerebrali”, atte a nutrire le larve.
A stadi diversi del loro ciclo vitale, queste attività sono prerogativa delle “operaie”, presenti in genere, specialmente negli alveari europei, con 50.000-100.000 unità, che si trasformano alla fine in “api bottinatrici” o “insetti pronubi”, deputati al complesso processo di raccolta del polline, produzione del miele e fecondazione dei fiori.
Mentre succhiano il dolce nettare di un fiore, raccolgono infatti, più o meno involontariamente, il polline con le “spazzole”, e lo trasportano con cestelli posti negli arti posteriori.
Le operaie sono inoltre dedite alla nutrizione e alle cure parentali della regina e delle larve nate dalle ovocellule, da lei deposte.
I fuchi, destinati solo alla riproduzione, oscillano tra i 500 e i 2000 per alveare.
Tutte le larve, sia quelle che genereranno operaie (le femmine), sia quelle da cui nascono i fuchi (i maschi), sia quelle che daranno luogo a una futura regina (femmina matriarca), hanno colore cereo e sono dette “eucefaliche” e “apode”, poiché la testa è molto sviluppata e sono prive di zampe.
Le larve destinate a formare le future regine si impupano dopo 15-16 giorni di vita; quelle che sono destinate a diventare operaie subiscono la prima metamorfosi a circa 21 giorni, mentre i maschi non si impupano prima di avere raggiunto il 24° giorno di vita.
Tutte le larve, indipendentemente dal morfotipo che produrranno, si convertono in una “pupa adectica”, cioè quasi del tutto immobile, “esarata o exarata” quindi con appendici libere rivestite di cuticola propria, ed “evoica”, perché la “crisalide” si rinchiude all’interno di un bozzolo di seta, o costituito da materiale prodotto dall’insetto in corso di sviluppo.
Tutte vivono allo stadio di pupa per un periodo che è diverso per ciascuna casta, quindi successivamente si trasformano in immagini (copia identica all’adulto), ed escono dalla loro cella dove erano state rinchiuse, sotto forma di “morfotipi alati”.
Ciò significa che la regina, l’operaia e il fuco, sono dotati d’ali rivestite di una membrana che s’indurisce al contatto con l’ aria, da cui il nome dell’ordine: Imenotteri (Hymenoptera), parola composta che significa “ali con membrane”.
La regina, accudita come detto dalle operaie, viene nutrita con “pappa reale” o “gelatina reale” (non avrà mai modifiche alimentari nel corso della sua vita), una miscela riccamente zuccherina ed energetica, anch’essa prodotta dalle “ghiandole sopracerebrali” delle operaie.
Le larve che daranno luogo ai fuchi ed alle operaie, sono, per un breve periodo iniziale, nutrite con la “pappa reale”, mentre successivamente con polline e nettare, sempre ovviamente solo dalle operaie nutrici. Un appellativo che ne identifica perfettamente l’efficienza e il ruolo.

L’evoluzione delle piante da fiore è strettamente legata alle api. Alcune, come questa salvia dei campi, hanno addirittura inventato dei poggia piedi e dei bilanceri per impollinare i pronubi © Giuseppe Mazza
La nutrizione, in termini quantitativi e qualitativi, agisce sulla maturazione dei caratteri sessuali secondari somatici, come il numero dei dentelli del pungiglione e le dimensioni dello stesso.
Le “ghiandole della cera”, dette appunto “ciripare”, presenti sempre solo nelle operaie, vengono invece utilizzate per costruire il favo, e chiudere l’ingresso della cellette esagonali con una pellicola cerosa che funge da porta.
All’interno di queste cellette, si può trovare una larva in sviluppo o incubazione con una certa quantità di cibo (pappa reale e poi anche polline se nasceranno operaie e fuchi), o solo cibo. In questo caso vengono usate come riserva di pappa reale, polline o miele, una volta terminata la trasformazione del polline da cui deriva.
La celletta esagonale è una caratteristica architettonica-strutturale interna dell’alveare, ripetuta in centinaia di copie (un vero modulo strutturale), all’interno della quale possono essere poste le larve neonate in incubazione, nel rapporto una celletta-una larva, o cibo come riserva alimentare.
La complessità geometrica, che si evidenzia osservando una sezione trasversale di alveare, in termini sia di numero di cellette, sia di organizzazione strutturale, che sfiora la geometria frattale, mostra l’elevato grado d’intelligenza sociale che caratterizza questi animali, a noi ancora così misteriosi, benché allevati da migliaia d’anni.
Una complessità simile, forse superiore, la si riscontra anche dai calchi ottenuti versando gesso liquido, dall’ingresso principale dei nidi vuoti di formiche, che pure appartengono all’ordine degli Imenotteri (Hymenoptera), e delle termiti, che fanno parte dell’ordine degli Isotteri (Isoptera) parola composta il cui significato è “ali con le medesime dimensioni”. Ne risulta sempre una costruzione complessa, comparabile ad una vera e propria architettura urbana, quasi una delle nostre città, con aree di riposo, aree di nutrizione, aree di riproduzione, strade, ecc.
Ritornando agli alveari, con le loro cellette chiuse, la larva in crescita vi troverà sempre il cibo fornito da un’operaia, mediante le ghiandole sopracerebrali, e un sufficiente volume d’aria per la respirazione durante lo sviluppo, mentre centinaia di operaie mantengono, fuori dall’uscio, la giusta temperatura e umidità dell’ambiente, parametri anch’essi fondamentali per il corretto sviluppo dell’insetto.

Il contratto con le piante è chiaro: io ti nutro e tu mi impollini © Giuseppe Mazza
In questa sorta di microincubatore a geometria esagonale, le larve restano fino allo “sfarfallamento”, quando esce la ninfa o immagine dell’ape, in tutto simile all’adulto, ma più piccola, che raggiungerà le stesse dimensioni dopo una serie consecutiva di accrescimenti.
In questo periodo, come accennato, avviene anche la matu- razione delle ali, chiamata “sclerificazione”.
Si asciugano dall’umidità che le ricopre e si irrobustiscono, rivestendosi di una membrana chitinosa-elastica, per essere atte al volo. Le varie venature caratteristiche, si organizzano in geometrie chiamate anch’esse “celle” e formano il sostegno dell’ala.
Da questo momento in poi la giovane ape si comporterà da operaia, se la larva è nata da una ovocellula fecondata con sperma di fuco (per anfigonia o amfimissi), o da fuco, se è nata da ovocellule non fecondate, mediante un processo partenogenetico, presente in questo imenottero domestico ed altre specie dell’ordine, detto “partenogenesi Arrenotoca”, senza fecondazione.
La regina mostra una totale mancanza di cure omoparentali verso lo sterminato numero di larve, formanti le numero- sissime generazioni di prole prodotte nel tempo. Si può definire una fredda “unità riproduttrice”, o meglio una “unità fattrice”, più che una madre, nel senso più letterario del termine.
È una vera e propria entità genica, con un cariotipo costituito da 32 cromosomi, devota alla propagazione dei suoi alleli e basta.
Tutte le cure dell’alveare, come mantenerlo pulito e adegua- tamente aerato, le cure parentali delle larve e delle ninfe, regina inclusa, toccano sempre alle numerosissime operaie, che in più, da autentiche stacanoviste, devono raccogliere il cibo, proteggere l’alveare dai nemici, e mantenere costante la temperatura interna e l’umidità del nido.
Devono provvedere insomma, a tutte le faccende essenziali perché la società in cui vivono, si mantenga in equilibrio omeostatico.
Abbiamo visto che i fuchi nascono per partenogenesi (ripro- duzione virginale Arrenotoca, quel tipo di partenogenesi che dà origine solo a individui di sesso maschile, contrapposta alla Telitoca, che genera solo individui femminili ed alla Deuterotoca, che dà origine, in proporzioni diverse, a individui di entrambi i sessi) dalle uova deposte dalla regina, che, essendo aploidi, hanno cariotipo n = 16 cromosomi.
Le femmine operaie, nate per anfigonia, ad opera della fusione gamica di uno spermatozoo con una ovocellula, come le nuove potenziali regine, hanno invece un cariotipo diploide 2n = 32 cromosomi.

Miele a parte, le api sono essenziali per le attività agricole umane © Giuseppe Mazza
Può eccezionalmente accadere che si producano maschi diploidi, per omozigosi. Queste larve vengono subito riconosciute dalle operaie nutrici e uccise.
Tale riconoscimento avviene forse per l’emissione da parte delle larve di sostanze feromoniche particolari, sesso-dipendenti, o per caratteri somatici (specifici della somazione), a noi ignoti, che non sfuggono alle nutrici.
Da queste numerose generazioni, nasce, prima o poi, una nuova regina diploide (2n), che sostituirà la vecchia, morta per malattia o uccisa dalle stesse operaie perché troppo anziana.
Ma in genere la vecchia regina continua a svolgere il suo ruolo, e la nuova migra altrove, con un certo numero di fuchi ed operaie, cercando un nuovo nido, per costituire una nuova società.
Tale fenomeno è detto “sciamatura”. In sostanza non sono presenti fenomeni di lotta territoriale per divenire capo della società in cui si è nati, ed esiste una forma latente di pace.
La “neo-regina” compie il volo nuziale a 5-6 giorni dallo “sfarfallamento”, accop- piandosi con un numero variabile (da 6-8) di fuchi, il cui sperma viene con- servato in apposite strutture dette “spermatofori”, dove si mantiene senza danni per settimane.
Lo userà, secondo un principio non ancora svelato (forse è in grado di comprendere, sia qualitativamente che quantitativamente, il rapporto numerico più adatto nella comunità tra operaie e fuchi), per fecondare le ovocellule rilasciate dai suoi “ovarioli”, che ormai ingrossati hanno gonfiato l’addome.
L’ovideposizione avviene dopo 1-3 giorni, per la nascita di operaie o, evitando la fecondazione, di fuchi o pecchioni aploidi, per partenogenesi Arrenotoca.
Spesso gli organi copulatori del maschio (fuco) vengono strappati per rimanere nella borsa copulatrice della femmina.

Una catena vivente d’operaie per la costruzione di un favo. Utilizzano la cera secreta da apposite ghiandole ©Mazza
Costituiscono quello che i biologi ento- mologi chiamano il “segno di fecon- dazione”, e verranno successivamente rimossi dalle operaie.
Questo vale anche per la vecchia regina, che come la neo-regina, migrata per sciamatura a creare una nuova società, compie parate nuziali accoppiandosi coi fuchi.
Le operaie sono più piccole sia della regina che dei fuchi, i veri pigri e nulla- facenti della società (giustificheremo poi il perché di questo). Possiedono le “ghiandole ciripare”, della cera, quelle “sopracerebrali” , ma sono fornite anche di cestelli e di spazzole nelle zampe posteriori, utili per la raccolta del polline quando vivranno come bottinatrici-pronubi.
Presentano anche loro ovarioli, ma in numero di 10-12. Sono quasi sempre inattivi o atrofizzati, sicuramente quando verso la vecchiaia si trasformano in bottinatrici; le operaie possono eccezio- nalmente essere chiamate a reggere il trono, quando la regina muore precocemente (per malattia o altra causa), e non è ancora nata una nuova regina pronta a rimpiazzarla.
Il processo è curioso. Una delle operaie, non è chiaro il criterio con cui avviene la scelta del soggetto su numeri dell’ordine di 50.000-100.000 unità, si trasforma (divenendo fertile) in regina “ad interim”, finché ne nasce una nuova, di ruolo, che prenderà il suo legittimo posto.
Le operaie, inoltre, sono le uniche api fornite di un pungiglione, o aculeo, connesso con una ghiandola velenosa, che provoca dolori lancinanti alla puntura.
Negli esseri umani, una puntura di Apis mellifera, come del resto accade col Calabrone asiatico (Vespa mandarinia), può provocare, quando il soggetto colpito è allergico, un grave “shock anafilattico”.
Se non viene soccorso subito, con farmaci cortisonici e antistaminici, il malcapitato rischia di morire, e questo vale anche per gli animali che vengono attaccati dalle api.

Oltre a costruire i favi, le operaie raccolgono il cibo, nutrono la regina, i maschi e le larve, mantengono costante la temperatura interna e l’umidità dell’alveare, che difendono, a costo della vita, da eventuali aggressori © G. Mazza
Mentre, dopo la puntura, l’aculeo può essere ritirato dai tessuti o dalla cuticola rigida
degli insetti, questo viene al contrario trattenuto, e strappato con le viscere, dalla pelle elastica dei mammiferi, provocando la morte dell’ape femmina. Un fenomeno più frequente nelle api che nelle vespe, dove c’è una maggior resistenza dell’aculeo saldato al corpo dell’insetto.
Prima ho definito i fuchi come i membri meno attivi della società, ma forse sono stato troppo severo nel giudicarli così negativamente.
È vero che il loro ozio apparente li fa sembrare dei perdigiorno, mentre ronza- no attorno all’alveare senza nemmeno curasi della difesa da eventuali nemici, ma non ne hanno colpa, perché ciò dipende dalla loro anatomia-fisiologia incompleta. Non sono dotati di un apparato boccale funzionante, e in ogni istante della loro breve vita dipendono quindi, per il cibo, dalle operaie.
Il ben noto meccanismo di ricompensa-fecondazione, che avviene quando un’ape bottinatrice-pronubo, succhia il nettare con la proboscide, dai nettàri floreali, è la molla dell’impollinazione dei fiori.
Fenomeno evolutivamente molto antico, indotto da condizioni ecologiche speci- fiche, che è alla base della riproduzione di una vasta gamma di piante, le Angio- sperme (Angiospermae), che si ri- producono mediante il “fiore”.
Mentre si nutre, passando di corolla in corolla, l’ape trasporta il polline raccolto da un altro fiore della medesima specie, questo è alla base della fecondazione incrociata.
Questo fenomeno riproduttivo, è stato definito dai biologi (botanici e zoologi), riproduzione “entomofila”, cioè per mezzo d’insetto.
Sarebbe lungo discutere di un meccanismo, che è a confine tra le due principali branche della Biologia (Zoologia e Botanica), e che ancora non ha svelato tutto ciò che ha da raccontarci.
Ad esempio, come e perché un’ape decida di visitare solo un determinato fiore, di una determinata specie vegetale, disinteressandosi invece di quelli di altre specie, o sul perché faccia ritorno più volte proprio su quel determinato fiore, per quella specifica specie vegetale (dimostrando capacità di memoria e discriminazione), disinteressandosi di fiori di piante conspecifiche vicine, che sono invece oggetto dell’interesse di altre bottinatrici del medesimo alveare e della medesima società.

Un telaietto estratto a mani nude dall’arnia. L’ Apis mellifera è molto meno aggressiva di quanto si creda © Mazza
Forse, in questo ultimo caso, avviene una sorta di marcatura individuo-specifica del fiore ospite, per cui non può più essere oggetto dei desideri di un altra compagna “pronubo”.
Questa caratteristica delle api, viene utilizzata in agricoltura biologica, per garantire la riproduzioni di piante eduli-officinali, come la colza, la zucchina, la zucca, le angurie e molte altre, ovviando alla fecondazione artificiale.
Da numerosissime ricerche etofisiolo- giche, entomofisiologiche e botaniche, di zoologia ed ecologia su campo, anche definita “fisiologia di campagna”, i biologi si stanno rendendo sempre più conto (a riguardo ci sono centinaia di pubbli- cazioni), che le piante da fiore, con la loro riproduzione entomofila, sono coevolute con le Api (Apis mellifera) e altre specie pronube.
I processi coevolutivi hanno fatto si che le piante che si affidano a questi pronubi, producano fiori in funzione degli occhi delle api, in grado di percepire la luce polarizzata ed i colori dello spettro, con lunghezze d’onda che cadono nel rosso-arancione-giallo.
È il motivo per cui poi troviamo spesso queste tinte nelle corolle, per non parlare degli “androcei” e “ginecei”.
L’occhio di un ape operaia è composto da circa 4000 ommatidi, mentre i fuchi, nei loro grandi occhi, ne contano più di 7500. Un chiaro carattere di dimorfismo sessuale.
Tali occhi complessi e compositi, permettono alle operaie, divenute “bottinatrici”, di discriminare tra le migliaia di fiori presenti in natura, quelli che hanno veri “néttari”, da quelli che non ce l’hanno.
È probabile che anche il profumo del fiore
giochi un ruolo specifico, fungendo da attrattore alimentare per l’ape, sempre al fine della riproduzione della pianta.

Un favo con le caratteristiche cellette esagonali. Larve a diversi stadi di sviluppo, polline e miele © Giuseppe Mazza
L’odore ha una forma convertibile in una variazione elettrica nel sistema nervoso dell’ape.
Quindi a un odore, percepito tramite le antenne, corrisponderà uno specifico “antennogramma”, che è l’equivalente fisiologico dell’elettroencefalogramma dei vertebrati.
È così che avviene il processo discrimi- natorio, di scelta e riconoscimento del fiore, da parte dell’ape, in base anche all’odore.
Stadi di sviluppo e corrispettivi funzionali
I vari compiti svolti dalle operaie, vengo- no suddivisi in relazione alla loro età.
Appena sfarfallate e con ali ancora non bene sclerificate, peregrinano tra i favi senza svolgere alcun lavoro, ma ben presto, divengono nutrici e balie sia della regina che delle larve in crescita ormai prossime a ninfosare.
Quindi, come scritto sopra, provvedono alla manutenzione delle cellette di cera, ripuliscono il nido e raccolgono il miele e il polline dalla compagne bottinatrici più anziane quando fanno ritorno all’alveare, una forma incompleta di “trofallassi”.
Dopo due o tre settimane divengono delle vere e proprie “amazzoni”, specializzate nella difesa dell’alveare, pronte a impedire l’ingresso a nemici, parassiti e persino ad api appartenenti ad
altre comunità.
Infine, ormai anziane, raggiungo il più alto grado della gerarchia a loro accessibile, divenendo appunto pronubi o bottinatrici.
Svolgono allora esclusivamente questo compito, volando anche a centinaia di metri dall’alveare, in cerca di fiori per lambirne il nettare con la proboscide e caricare il polline.
Questo viene ingurgitato in un particolare diverticolo dell’esofago, la “tasca mielaria”, dove è subito sottoposto a processi di digestione enzimatica ad opera di glicosidasi, a cui farà seguito, nel nido, l’evaporazione di parte del contenuto acqueo, per arrivare alla formazione del dolce prodotto che tutti conosciamo: il “miele”.

Uovo appena deposto in posizione verticale. Due uova orizzontali anziane ed una che si trasforma in larva © Mazza
Per essere più precisi, non tutto il polline finisce nella “tasca mielaria” per la dige- stione: una parte viene semplicemente ammassata sulle spazzole, nelle zampe posteriori e trasferita, per mezzo dei pettini, nei cestelli delle tibie per il tra- sporto al nido.
Qui viene immagazzinato come riserva di cibo, o convertito in miele all’interno delle cellette.
A fine giornata, le bottinatrici stanche morte, fanno ritorno all’alveare per risposarsi.
Il linguaggio delle Api
Nella società delle api, come in quelle degli altri insetti eusociali, esiste un linguaggio più o meno codificato, che può essere più o meno mimato.
Ad esempio, quando nasce una regina, pronta a migrare dal vecchio nido per formare una nuova società, un gruppetto di operaie e fuchi la precedono per cercare la sede idonea, come l’interno di un albero cavo o un anfratto roccioso.
Quando hanno trovato il posto adatto, ne segnalano l’ubicazione alla regina con codici simulati, mediante movimento del corpo e modalità di volo. Questa capisce e spicca solennemente in volo, seguita da tutto il suo popolo.
Nel nuovo nido le operaie, secerneranno la cera e iniziano la costruzione dei favi, costituiti dalle classiche cellette esagonali, mentre la regina, dopo essersi accoppiata coi fuchi, accudita e difesa, dalle altre api femmine, inizia a deporre migliaia e migliaia di uova, fino alla fine della sua lunga vita, che può raggiungere i 4 anni, un durata enorme, nel mondo degli insetti.
Ma le prove più complete e concrete sul linguaggio, mimato in danza, dalle api, furono ottenute dagli storici e sensazionali esperimenti compiuti per 30 anni, dal biologo austriaco Karl von Frisch, che vinse, in collaborazione con altri due biologi, Konrad Lorenz e Nikoolas Tinbergen, il premio
Nobel per i suoi studi nel 1973.

Le larve, dal colore cereo, hanno la testa molto sviluppata e sono prive di zampe © Giuseppe Mazza
Col biologo tedesco Oskar Heinrot, questi tre scienziati sono considerati i fondatori dell’Etologia animale scientifica.
Come abbiamo descritto sopra, le api sono dotate di un complicato linguaggio, che utilizza oltre alla chemiorecezione, stimoli tattili, visivi e uditivi.
K. von Frisch e colleghi, hanno compiuto accurate osservazioni sui mezzi di comu- nicazione delle api e hanno scoperto almeno due fondamentali meccanismi usati da questi imenotteri sociali, per
informare i compagni di alveare (più precisamente le compagne) sull’ubi- cazione del nettare, del polline e di altre sorgenti alimentari.
Una “danza circolare” delle bottinatrici, andate in esplorazione, indica che il cibo trovato è situato a poca distanza dal favo, mentre una detta “danza della coda” (eseguita in parte dimenando l’addome, cioè “scodinzolando”) segnala che il cibo è lontano.
Di solito ambedue le danze vengono eseguite all’interno dell’alveare, al buio, per cui solo tramite i chemiocettori e i recettori tattili situati sulle antenne, che vengono poggiate sulle bottinatrici danzanti, le altre api operaie possono rendersi conto di quale tipo di cibo si tratta e, seguendo dettagliatamente la forma della danza, possono ricavare informazioni sulla distanza e sull’ubicazione della sorgente alimentare. Talvolta una sommesso e particolare tipo di ronzio accompagna questa danza.
Le api bottinatrici danzanti, reduci da una raccolta di alimenti, dopo aver ripetuto la danza più volte, tornano in volo verso la sorgente di cibo, accompagnate da altre api pronube, che hanno compreso l’informazione ricevuta.
Finché la sorgente alimentare rimane cospicuamente abbondante, la danza viene ripetuta ad ogni ritorno al favo; ma quando la risorsa alimentare è in fase di esaurimento, le api bottinatrici, continuano a trasportare il cibo, ma ad ogni ritorno non compiono più la danza informatrice.

I fuchi servono alla fecondazione della regina. Per l’apparato boccale incompleto sono nutriti dalle operaie © Mazza
Infatti non è più necessario aumentare il numero delle bottinatrici per trasportare il cibo.
La “danza circolare” esplicata quando la fonte di cibo è vicina all’alveare, consiste in una serie di piccoli cerchi, prima in una direzione (ad esempio senso orario o viceversa) e poi nella direzione opposta (senso antiorario o viceversa).
Quando le api esploratrici, hanno ripetuto la danza più volte, lasciano l’alveare, seguite da altre bottinatrici eccitate dalla danza informativa.
Voleranno intorno all’alveare, finché localizzano la fonte di cibo e vi si dirigono.
La “danza della coda”, usata quando la fonte del cibo è lontana, per dare la direzione e la distanza della sorgente alimentare, viene compiuta su una superficie verticale dell’alveare, e consiste di tre fasi distinte: una passeggiata semicircolare, un volo in avanti, e una nuova passeggiata semicircolare, in direzione opposta alla precedente, che completa un cerchio immaginario.
La direzione della sorgente di cibo, rispetto all’alveare, è indicata dal breve volo in avanti: se questo è verticale verso l’alto, il tragitto da compiere è in direzione del sole; se è verticale verso il basso, il tragitto è in direzione opposta al sole; se è in diagonale, il tragitto è spostato di qualche grado d’angolo rispetto la posizione del sole.
Il ritmo, con cui viene condotta tale danza, indica la distanza del luogo in questione rispetto l’alveare.
Un ritmo lento, di circa 4 danze al minuto, indica una distanza di circa 10 km; 16 danze per minuto, indicano una distanza di circa 1 km. Esiste quindi una relazione di inversa proporzionalità, tra il ritmo della danza e la distanza da percorrere: maggiore è il ritmo della danza, minore la distanza da percorrere e viceversa.
Altri dettagli della danza (la sua persistenza nel tempo, il numero degli scodinzolii durante il volo in avanti, il ronzio che li accompagna, ecc.) forniscono ulteriori informazioni utili, alle colleghe bottinatrici.
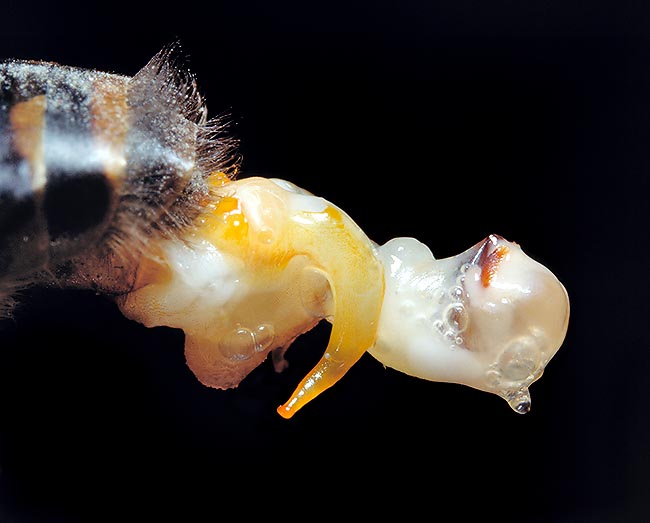
Apparato sessuale di un fuco. La regina si accoppia con 6-8 fuchi e conserva lo sperma per settimane © G. Mazza
Ciò dimostra come questo complesso lin- guaggio sia uno dei più ricchi nel mondo degli insetti.
Per comprenderlo fino in fondo, molto deve essere ancora spiegato.
Una sua chiarificazione, può essere utile sia per una più razionale e sana agricoltura e forse, potrà spiegarci perché negli ultimi 10 anni, in tutta Europa e non solo, si sta assistendo a fenomeni di decadimento delle popolazioni di Apis mellifera, affiancati da un ridotto indice riprodut- tivo.
Apicoltura e parassiti
Prima di parlare del grande ruolo svolto dall’ape domestica (Apis mellifera) nella produzione del miele e della cera, oltre che della pappa reale e del “propoli” (sostanza resinosa raccolta dalle operaie da varie specie di piante), attività oggi organizzate in una vera e propria filiera industriale, che va sotto il nome di “Apicoltura o Apicultura” (e chi la svolge è detto apicoltore o apicultore), dobbiamo anzitutto brevemente accen- nare alle varie specie e razze di api note alla Biologia.
Il genere Apis comprende fondamentalmente quattro specie.
La prima è quella di cui abbiamo fin qui parlato: l’ Apis mellifera.
Una seconda specie è l’Ape gigante (Apis dorsata) dell’Asia meridionale, difficile d’allevare perché molto aggressiva. Può infatti aggredire, con grandi sciami, animali ed uomini, a volte con effetti letali.
Una terza specie è l’Ape indiana (Apis indica), diffusa dall’India all’Africa, caratterizzata da scarsa capacità di lavoro e tendenzialmente nomade, poco interessante quindi ai fini dell’allevamento.

Favo con cella di ape regina © Giuseppe Mazza
La quarta specie è l’Ape nana (Apis florea o Apis cerana), che predilige le regioni più calde, a bassa quota, dell’Asia meridionale.
Vive in società di piccole dimensioni, e alveari modesti, produ- cendo solo piccole quantità di miele, più o meno equivalenti a quelle prodotte dalle vespe, e quindi del tutto irrilevanti dal punto di vista commerciale.
L’ Apis mellifera è a sua volta suddi- visa in diverse razze o sottospecie, come l’ Apis mellifera ligustica, che è una delle più note, scoperta negli anni ’30 dai biologi entomologi, e l’ Apis mellifera mellifera.
Non mancano infine ibridi artificiali, che si sono introdotti negli ecosistemi naturali.
È il caso della ben nota Ape Africanizzata o Ape killer, prodotta involontariamente dal biologo brasiliano DrSc Warwick Estevam Kerr.
Il DrSc W.E. Kerr, esperto di api, stava compiendo esperimenti nel dipartimento di entomologia della facoltà di Scienze Biologiche dell’Università di Brasilia, per ottenere api più attive, e quindi più produttrici di miele, in ambienti tropicali-caldi, come quello Brasiliano.
Il suo scopo, ovviamente, era di soste- nere l’apicoltura locale.
Alla fine degli anni ’50, aveva introdotto alveari di api della Tanzania, in partico- lare della razza africana Apis mellifera scutella. Api più piccole di quelle europee (Apis mellifera e le relative razze) e più aggressive, ma più idonee agli ambienti tropicali.
Involontariamente, qualche decina di regine di Apis mellifera scutella, fuggirono dagli alveari e
furono fecondate dai fuchi delle razze Apis mellifera ligustica e Apis mellifera mellifera, fino allora usate, anche in Sud America, dagli apicoltori.
Ne derivò un ibrido biologico aggressivo, come l’ Apis mellifera scutella, ma di maggiori dimensioni, e quindi più pericoloso della sottospecie o razza originale.
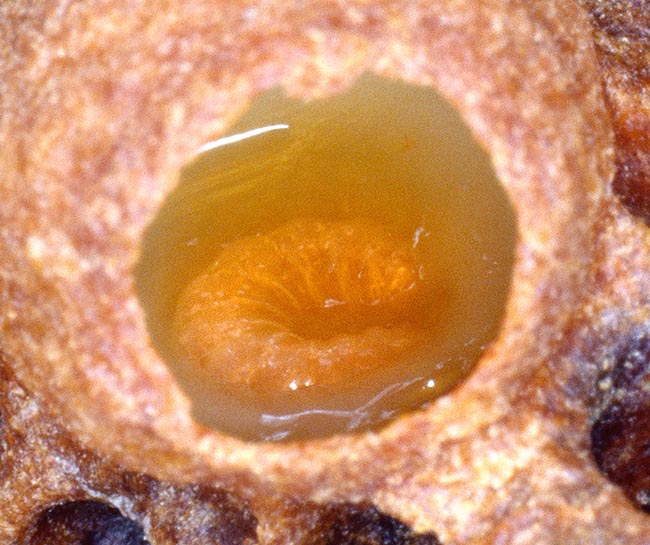
Larva di regina nella sua cella. Sarà nutrita per tutta la vita dalle operaie con pappa reale © Giuseppe Mazza
L’allevamento delle api, di preferenza Apis mellifera, praticato per ottenere miele, propoli, cera, pappa reale, sciami e regine ha origini antichissime.
Queste risalgono alle epoche degli Egizi, dei Babilonesi e quindi dei Romani.
Attività tenuta in grande considerazione quando il miele era l’unica sostanza dolcificante conosciuta, che decadde con la scoperta dell’America e la conseguente diffusione dello zucchero (da canna e barbabietola), per non parlare della diffusione di materie sostitutive della cera.
Alla vita delle api, ed al loro razionale sfruttamento, vennero dedicate numero- se opere di poeti e di scrittori georgici.
Con l’adozione dell’arnia a favo mobile, dei fogli cerei stampati e dello “smelatore centrifugo”, si può dire che ebbe inizio la moderna apicoltura, che non richiede l’apicidio per estrarre il miele.
Ancora oggi i Kikuyo, popolazione tribale degli altipiani della Tanzania e del Kenya, golosi del miele aromatico prodotto dalle api autoctone, quando ne vanno a caccia, utilizzano la fumigazione per confondere le operaie a difesa del favo, e lo distrug- gono per estrarre il miele, incuranti delle innumerevoli punture.
Secondo alcuni apicoltori europei, tale miele sembrerebbe più aromatico e gustoso di quello prodotto nel Vecchio Continente.
Il successo dell’apicoltura è molto legato alla scelta della località d’impianto dell’apiario, alle colture che vi si praticano e all’andamento delle stagioni.
Per assicurare l’alimentazione delle api ed una buona produzione di miele, non è sufficiente la fioritura primaverile. Occorre che questa si protragga fino all’estate e possibilmente all’autunno, condizioni che in genere si realizzano in località situate a medie altitudini.

L’ape regina marcata in azzuzzo viene nutrita per trofallasi da un’operaia. Può vivere circa 4 anni, durata enorme nel vasto mondo degli insetti. Per riconoscerla subito nell’alveare viene marcata alla nascita dall’apicoltore con una vernice che asciuga all’istante. I colori ammessi sono cinque, con un preciso calendario internazionale. Così il 2010 e il 2015 è, come nella foto, l’anno dell’azzurro, il 2011 e il 2016 l’anno del bianco, il 2012 e il 2017 quello del giallo, il 2013 e 2018 del rosso, e il 2014 e 2019 del verde © Giuseppe Mazza
Precedendo da Nord a Sud, il pascolo ideale è rappresentato dai prati di trifoglio, dai frutteti (rosacee), dal tiglio, dall’erba medica, dagli agrumeti, dai carrubeti e dai sullai.
Le attrezzature necessarie, non sono costose, salvo che si voglia procedere a grossi impianti a scopo industriale.
L’incremento dell’allevamento apistico, è sem- pre stato connesso alla diffusione delle associazioni di apicoltori, e la legislazione in materia, ha posto quindi, a fondamento di questa disciplina, la costituzione di appositi consorzi, che possono essere facoltativi o obbligatori.
Finiamo accennando brevemente ad alcune patologie parassitarie, che sono da sempre causa di malattie per le api, fino a determinare la morte di intere colonie, per non parlare dell’attuale sconvolgimento delle condizioni ambientali, e dell’inquinamento elettromagne- tico “elettrosmog”, causato dalla sempre più pressante e veloce urbanizzazione, che portano varie specie di api a un vero e proprio collasso, compromettendone anche le capacità riprodut- tive.
I biologi entomologi da campo, avevano già alla fine del secolo XVIII° e inizio del XIX°, individuato alcuni parassiti infestanti e letali per le api, ma con lo sviluppo dell’apicoltura, i biologi hanno potuto chiarirne più in dettaglio i cicli vitali e parassitari.
Tra i vari parassiti, due sono dei veri e propri flagelli per le colonie di Apis mellifera, con conseguenti danni commerciali.
Si tratta dell’acaro ectoparassita (Varroa destructor) e dell’Acaro tracheale (Acarapis woodi).
Questi acari parassiti vengono facilmente trasmessi dalle regine durante la “sciamatura”, con i fuchi e le operaie al seguito, o durante il volo di stock di api, alla ricerca di nuovi nidi.
Quando tali agenti si insediano in un favo, causano inevitabilmente l’alterazione distrofica dell’omeostasi coloniale, portandola alla decadenza totale, fino all’esaurimento completo.
Ciò sta creando seri problemi all’apicoltura e quindi all’industria del miele, ma il rischio principale, secondo i biologi, è che questi parassiti, in associazione con altri fenomeni dannosi-ambientali, possano portare all’estinzione dell’ Apis mellifera,
con danni incalcolabili sull’equilibrio ecologico globale.

La regina mentre depone le uova: 2.000 al giorno e qualche milione nell’arco della vita © Mazza
Il Varroa destructor, come sottolinea il nome, è particolarmente distruttivo per questa specie: il principale agente biologico responsabile del calo demografico dell’ Apis mellifera, con enormi danni per gli allevamenti.
Si è visto che le api colpite da questo agente, modificano, anticipandola, la fase del foraggiamento, perdono le capacità di volo e orientamento, e si verifica un rapido aumento della mortalità invernale, sia delle singole unità che per le intere colonie, poiché è il parassita si diffonde facilmente all’interno del favo, quando, per esempio, una operaia infestata vi fa ritorno.
L’Acaro tracheale (Acarapis woodi) è una malattia infestante a impatto più variabile.
Diversi studi di biologi entomologi britannici degli anni ’70, hanno dimostrato che esso potrebbe non essere un’attuale minaccia per la stabilità demografica della specie Apis mellifera, mentre fu la causa biologica delle devastazioni avvenute negli Stati Uniti setten- trionali durante il XVIII°secolo.
Questi studi hanno anche dimostrato che l’ Acarapis woodi risulta molto più efficiente, e quindi patogena, nei paesi freddi, come negli Stati Uniti del Nord e in Nord Europa per l’emisfero Boreale, o all’estremo Sud, per quello Australe.
Una volta insediatosi in una colonia di Apis mellifera, l’acaro tracheale, determina tendenzialmente una riduzione dell’indice riproduttivo, una difficoltà d’allevamento delle larve, da parte delle
operaie, e del loro impupamento, con un incremento della mortalità invernale, associata ad una più alta incidenza infettiva virale e batterica.
Se non viene trattato tempestivamente dall’apicoltore, il Varroa destructor, che ricordiamo essere un acaro ectoparassita, è ancora più virulento. In appena due anni può far collassare, distruggendola, una intera colonia di api domestiche.
Il Varroa destructor, è un ectoparassita abbastanza grande, che si sviluppa e cresce nella cella dell’ape in sviluppo.

Quando nasce una nuova regina, al centro a destra della foto, sciama con le operaie e i fuchi per fondare una nuova colonia © Mazza
Le femmine adulte di questo acaro, entrano nella cella appena prima che la larva si impupi, rilasciano le uova e pungo l’ape in sviluppo, per suggere l’emolinfa di cui si nutrono,come del resto le uova in sviluppo.
Il Varroa destructor e la sua prole, non uccidono l’ape in sviluppo (vero parassitismo). Sopravvivono durante il periodo di sviluppo (foretico) dell’ape, e poi si attaccano alla ninfa, fino a poco prima lo sfarfallamento, per passare in fine ad un’altra cella, con una nuova larva in sviluppo.
L’Acaro tracheale (Acarapis woodi) svolge invece il suo ciclo vitale completo nelle trachèole dell’ospite adulto.

Un apicoltore intento al suo lavoro. Un alveare può produrre anche 20 kg di miele all’anno © Giuseppe Mazza
Negli insetti non esistono infatti dei pol- moni, e si parla di specie “pneustiche”, che respirano con le trachèole, in contrapposizione alle “apneustiche” che, prive di trachèole, respirano mediante la cute.
Pungendo le trachèole, succhiano l’emo- linfa, finché l’ospite (un ape adulta mas- chio o femmina) non muore.
La proge- nie lascia quindi l’ospite, solo per trovare un’altra povera ape suscettibile all’infe- stazione.
In passato i biologi credevano che un alveare potesse essere attaccato, volta per volta, da uno o dall’altro parassita, ma ricerche recenti, su campo, hanno provato che nei favi di una colonia possono essere presenti entrambi i parassiti letali.
Gli studi dei biologi da campo, sulla Biologia evolutiva ospite-parassita e quelli sulla cosiddetta Immunoecologia, sono oggi fondamentali per dipanare la complicata matassa che regola questi processi d’infestazione, e trovare una cura efficace, non ancora disponibile, a salvaguardia dell’ Apis mellifera, che, non dimentichiamolo, gioca anche un ruolo importante nell’impollinazione di molte specie vegetali, ed è senza dubbio “essenziale” all’equilibrio e al manteni- mento della “biosfera”.