Familia : Ornithorhynchidae
Texto © Claudio Lanzoni

Traducción en español de Beatrice Marini

El pico del Ornithorhynchus anatinus posee muchas terminaciones nerviosas © Giuseppe Mazza
Características morfológicas, anatómicas y fisiológicas
El ornitorrinco tiene un cuerpo regordete y plano y un pelaje espeso e impermeable, de un color marrón intenso en la espalda y amarillento en el vientre, en todo el cuerpo excepto en las patas y en el pico.
El pelaje cubre también los ojos y las orejas (que están desprovistas de pabellón auricular).
La cola está compuesta especialmente por grasas que utiliza como reserva energética durante el invierno o en tiempos de escasez de alimentos. Puede llegar a medir hasta 10-15 cm de largo y es parecida a la del castor, aunque tiene forma y función diferentes. En efecto, la cola del castor, ancha y plana, es empleada para avanzar en el agua, mientras que el ornitorrinco, que utiliza las patas anteriores para propulsarse, sólo usa la cola para maniobrar en el agua.
Las patas son cortas, con 5 dedos provistos de garras, y las anteriores son palmeadas. Su andadura en la tierra firme es reptiliana (las patas están a los lados del cuerpo).
Mediamente los machos adultos son un tercio más grandes que las hembras (unos 50 cm de largo y 2 kg de peso). Ellos presentan, en ambas las patas posteriores, y más precisamente en el tobillo, un espolón córneo, que mide apróximadamente 1,5 cm. Este espolón es hueco y está conectado, a través de un conducto, a una glándula venenosa. Las hembras jóvenes poseen una versión rudimentaria de esta glándula, pero la pierden en los primeros años de vida. La equidna también tiene un espolón rudimentario y algunas glándulas venenosas que ya no funcionan.
El pico del ornitorrinco, provisto de dos narices, es bastante flexible (parecido al del pato), suave, gomoso y muy sensible al tacto. Esta característica se debe a la presencia de numerosas terminaciones nerviosas. El animal utiliza el pico para buscar alimento y orientarse en el agua. Este animal es desprovisto de dientes (que todavía son presentes en los recién nacidos) y trituran los alimentos gracias a algunas placas afiladas que están dentro de la boca.
El ornitorrinco posee un sistema sensorial especializado para percibir la presencia de presas en el agua. Este sistema se basa en la relevación de campos eléctricos generados por las presas. El pico está provisto de electrorreceptores situados en hileras rostrocaudales y mecanorreceptores distribuidos uniformemente por el pico. La zona electrosensible de la corteza cerebral se encuentra en el área somatosensorial táctil y algunas células corticales reciben estímulos tanto de los electrorreceptores como de los mecanorreceptores, lo que sugiere una fuerte asociación entre los sentidos del tacto y de la electrorrecepción.

Las patas anteriores son iguales en ambos sexos © Giuseppe Mazza
Como explicado antes, cuando el ornitorrinco busca alimento, ello utiliza prevalentemente su pico, con lo que excava en el fondo de los riachuelos. Es posible que los electrorreceptores se utilicen también para distinguir los objetos “animados” de los “inanimados”.
Sin embargo, el sistema de electrolocalización de los ornitorrincos presenta muchos lados obscuros y necesitaría investigaciones más profundizadas. Respecto a otros mamíferos tiene un metabolismo más bien bajo y su temperatura corporal media es inferior a la de los demás (32o C).
Ecología y comportamiento
Los ornitorrincos son animales nocturnos y semiacuáticos, nadadores excelentes, que viven en ríos, lagos y riachuelos de casi toda Australia y Tasmania, aunque están mucho más presentes en el sudeste del continente australiano, donde las llenas de los ríos (que causan daños a sus nidos) son menos frecuentes y la presencia de cocodrilos (por los que llegan a ser presas fáciles) es más rara.
Es un animal más bien solitario, que vive y se alimenta en territorios particulares. El territorio del ornitorrinco puede coincidir con el de otros, pero al parecer no subsisten peleas para controlarlo. El ornitorrinco crea dos diferentes tipos de madrigueras: una madriguera para vivir y una madriguera destinada a la reproducción. La primera es utilizada tanto por el macho como por la hembra para alimentarse y descansar, la segunda, más compleja, la utilizan las hembras para criar a los recién nacidos.
Las madrigueras para la reproducción pueden medir hasta 7 metros de altura y 18 metros de largo con entradas por encima y debajo del agua . En la temporada de apareamiento, la hembra cierra las entradas de la madriguera cada vez que entra o sale de ella, y las crías se quedan en el nido por unos 3 meses. La entrada por encima del agua serviría tanto para favorecer la entrada de oxígeno como para proteger a las crías de la subida del nivel del agua.
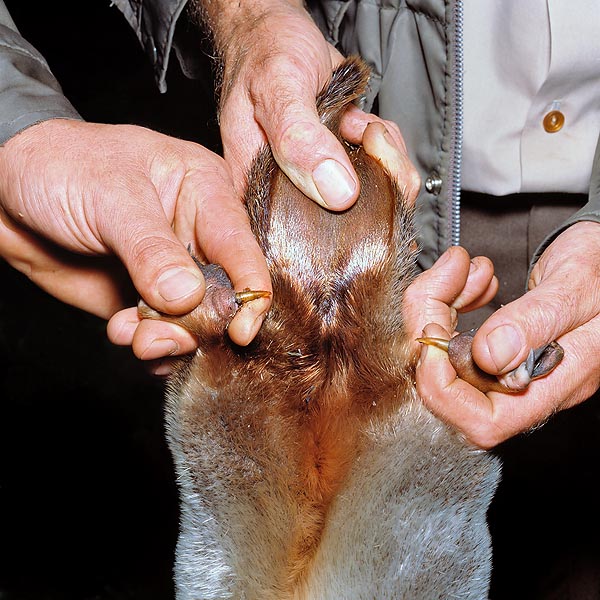
Las patas posteriores del macho poseen un espolón venenoso más © Giuseppe Mazza
Por la noche el ornitorrinco sale a buscar su comida.
Es carnívoro y se alimenta de invertebrados (crustáceos, lombrices, insectos), huevos de peces y anfibios, y pequeños vertebrados (peces, ranas, renacuajos).
Durante el verano se alimenta mucho más abundantemente que en invierno, acumulando las reservas en exceso en la cola, en forma de grasa. La busqueda de alimento occurre durante sus inmersiones en el fondo de los ríos, utilizando la cabeza y el hocico para desplazar el fango.
Sus inmersiones duran unos 40 segundos y, durante este periodo, el ornitorrinco utiliza, para localizar a sus presas, tanto el tacto como su capacidad de percibir las variaciones eléctricas (gracias a las terminaciones nerviosas de su pico, del que hemos hablado precedentemente).
Una vez capturada la presa, el ornitorrinco la desplaza en el saco maxilar, que está situado detrás del pico. Pues el ornitorrinco asciende a la superficie y la desplaza en la boca, donde la tritura con sus placas, situadas en la mandíbula. A los lados de la mandíbula hay puntas denticuladas que utiliza para eliminar los residuos (cáscaras, fango, etc.).
Reproducción
Los ornitorrincos alcanzan la madurez sexual alrededor de los dos años. En la etapa precedente de sus vidas, los machos no producen esperma y las hembras ocasionalmente ponen huevos. La temporada de apareamiento tiene lugar entre julio y agosto. El cortejo empieza en el agua. Normalmente la hembra toma la iniciativa, los dos nadan juntos y se aparean en el agua. El periodo de gestación es de 2-3 semanas. La hembra no da luz a crías vivas, sino pone huevos en un nido. Los huevos son normalmente dos y son incubados durante alrededor de diez días por la hembra en el fondo de la madriguera.
Las crías, sin pelo, miden unos 18 mm; ellas se quedan en la madriguera, alimentándose con la leche de la madre por 3 o 4 meses. Una vez que salen del nido siguen nutriéndose con la leche por un determinado periodo, hasta que puedan buscar alimento autónomamente. El cuerpo de la hembra produce una leche particular (desprovista de lactosa).
La leche se suministra a las crías a través de dos estructuras, parecidas a pezones, situadas en el abdomen y rodeadas por el pelo. Los recién nacidos, comprimiendo estas zonas, provocan la salida de la “leche”, que gotea del pelo, y pueden así lamerla.
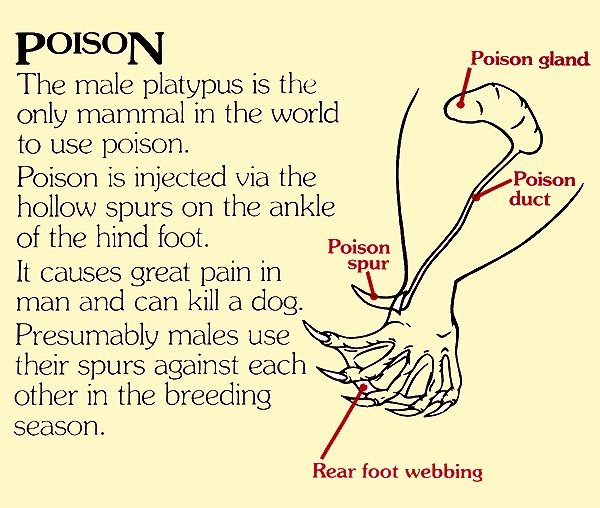
Como muestra este panel del Healesville Sanctuary el espolón está conectado con una glándula venenosa
El analisis de la genética del ornitorrinco también ha demonstrado que este animal es una mezcla de caracteristicas más complejas de lo que se podría creer (Grutzner et al., 2003).
Se teoriza que la especie se separó de su antepasado que tenía en común con la especie humana, hace unos 170 miliones de años. Recientemente se ha descubierto que el ornitorrinco posee 10 cromosomas sexuales (10 X en la hembra y 5X y 5Y en el macho) (Waters et al,. 2005).
Los científicos han buscado en el genoma del Ornithorhynchus tanto las secuencias de DNA pertenecientes a los Monotremas como las que están presentes en otros Vertebrados, descubriendo que algunas secuencias genéticas se parecen más a las de las Aves que a las de otros Mamíferos (Grutzner & Graves, 2004, Rens et al., 2007).
Por ejemplo, el cromosoma DMRT1, desde hace mucho tiempo estudiado por los genetistas porque parece determinar el sexo de las aves (sistema WZ), se encuentra casi igual también en el ornitorrinco (Veyrunes et al., 2008).
Estos elementos hacen pensar que la aparición de los sexos no ha sucedido separadamente en las aves y en los mamíferos, sino que una parte de esta diferenciación es común.
El aparato venenoso del Ornitorrinco: estructura y funciones
El aparato venenoso o sistema crural de los ornitorrincos sólo se refiere a los machos. Este consta de un par de espolones corneos vacíos, uno por cada pata posterior, y están localizados cerca del tobillo. Cada espolón está rodeado por la musculatura y está fijado a una articulación que le permite de quedarse erguido en caso de necesidad. A la base del espolón está situado un tanque del que parte un conducto delgado. El canal central del espolón de un ornitorrinco macho y adulto tiene un diámetro de unos 0,2 mm (Whittington et al., 2009).
Largos conductos, a través de los bíceps, conectan los tanques con las glándulas venenosas (alveolares, con forma de riñón). Cuando ataca a un rival, el macho erige sus espolones y los planta en la carne de la víctima; le inócula el veneno a través de inyecciones repetidas y, durante cada ataque, puede inyectar hasta 4 ml de veneno (Whittington et al., 2009). En el pasado han sido muchas las hipótesis sobre la función del espolón y de las secreciones emitidas por las glándulas femorales.

Su andadura en la tierra firme es cohibida y reptiliana © Giuseppe Mazza
Los espolones pueden ser utilizados con finalidad de defensa para contrastar a predatores potenciales (como cocodrilos, rapaces, demonios de Tasmania) pero, durante la temporada de apareamiento, representan también un excelente sistema de ofensa para vencer a los otros machos de la misma especie. La hipótesis dominante, ya formulada por Martin y Tidswell (1895) es que, siendo presente sólo en los machos adultos, los espolones deben de ser conectados con operaciones de selección sexual.
La constatación de que el ornitorrinco tiene pocos enemigos representa la demonstración que existe una relación entre la actividad secretoria de las glándulas femorales y la reproducción; en efecto, durante la temporada de apareamiento la glándula del veneno aumenta de dimensión, crece la cantidad de veneno producido y aumentan las dimensiones del testículo (Temple-Smith, 1973; Grant y Temple-Smith, 1998; Torres y Kuchel, 2000): esto avala la idea que el sistema crural haya evolucionado para tener una función reproductiva.
El envenenamiento de los rivales otorgaría una ventaja a los machos dominantes: la inyección del veneno, causando dolor e impediendo los movimientos, permite al macho dominante aparearse sin obstáculos y sin necesidad de luchar. El envenenamiento de un ornitorrinco por parte de otro macho normalmente no es fatal. Todavía las manifestaciones clínicas del envenenamiento han sido poco estudiadas.
Los primeros casos documentados de envenenamiento humano se remontan al final de 1800 (Martin y Tidswell, 1895), pero la descripción del primer caso clínico sólo se remonta al 1992 (Fenner et al., 1992). En este trabajo se muestran los principales síntomas del envenenamiento que son edema e inmediato, agudo dolor que ni siquiera puede ser calmado con morfina y persiste durante meses (Fenner et al., 1992). Las pruebas hematológicas realizadas en laboratorio muestran un aumento de la velocidad de eritrosedimentación, que indica una probable coagulopatía; de todos modos no parece que exista evidencia de neurotoxicidad, miotoxicidad o efectos necrotizantes. No han sido documentados casos mortales que incluyan al ser humano. Síntomas similares (dolor y edema) han sido descritos también por autores sucesivos (Tonkin y Negrine, 1994).

En cambio en el agua es muy ágil y percibe los campos eléctricos generados por las presas © G. Mazza
La información que tenemos sobre las substancias que componen el veneno del platipo todavía es muy escasa. Son pocos los estudios llevados a cabo en este campo hasta ahora, y es difícil realizar adicionales investigaciones debido a los problemas de conseguimiento del veneno y muestras de tejido.
El veneno del ornitorrinco es una sustancia más bien compleja. La inusual naturaleza de los síntomas daría la idea de una mezcla compleja de muchos diferentes péptidos y proteínas, algunas todavía desconocidas (de Plater et al., 1995; Whittington y Belov, 2007).
Los primeros experimentos conocidos sobre la secreción de las glándulas crurales han demonstrado que el veneno provoca, en los conejos, un grave edema y una rápida disminución de la presión arterial (PA) a través de una inyección subcutánea y dificultades respiratorias y muerte si inyectado por vía venosa. En los años ’90, la disponibilidad de nuevas técnicas para la caracterización de las proteínas ha permitido identificar con precisión algunos componentes del veneno del ornitorrinco. Algunos estudios han demonstrado las consecuencias del veneno en la musculatura lisa, permitiendo ipotizar la presencia al interior de un péptido natriurético (de Plater et al., 1995; 1998 a,b). Los péptidos natriuréticos son una familia de péptidos que se han conservado en las diferentes especies y que parecen controlar la presión sanguínea.
Recientemente se ha descubierto que el veneno consiste al menos en 19 diferentes fracciones peptídicas (Whittington y Belov, 2007) junto con componentes no proteicos (probablemente prostaglandinas). Hoy en día se han identificado sólo tres péptidos y son secuenciales aunque sus funciones específicas todavía no se conocen con precisión. Ellos son: péptidos natriuréticos de tipo C (CNPs); defensin-like peptides (DLPs); nerve growth factor (NGFs).
Sinónimos: Ornithorhynchus brevirostris Ogilby, 1832; Ornithorhynchus crispus MacGillivray, 1827; Ornithorhynchus fuscus Péron, 1807; Ornithorhynchus laevis MacGillivray, 1827; Ornithorhynchus novaehollandiae Lacépède, 1800;
Ornithorhynchus paradoxus Blumenbach, 1800; Ornithorhynchus phoxinus Thomas, 1923; Ornithorhynchus rufus Péron, 1807; Ornithorhynchus triton Thomas, 1923.